основные характеристики и особенности их применения
ГлавнаяРазноеМембрана диффузионная
Диффузионная мембрана (пленка): Что это такое
Диффузионная мембрана — это рулонный материал для проведения гидроизоляции. Его используют для наружных работ и он является важным звеном в утеплении дома. Материал характеризуется односторонней проводимостью влаги и способен удалять пар из утеплителя, который образуется из-за перепадов температур и существенного повышения влажности.
Строение диффузионной мембраны
Она обычно трехслойная, верхний и нижний слой — это полотна из нетканого полипропилена. В мембранах Ондутис средний слой устойчив к растяжениям. Вместе с крепким двухслойным каркасом мембрана характеризуется высокой стойкостью к механическим повреждениям, гидрофобностью и паропроницаемостью. Слои в диффузионных мембранах соединяются с помощью ультразвука — это исключает возможность повреждения середины и позволяет достичь отличных параметров. Такие мембраны экологически безопасны и не подвержены пагубному воздействию бактерий.
Паропроницаемость — важный параметр диффузионной мембраны. По этой характеристике материал делится на три категории:
- Мембраны малой диффузии материал, паропроницаемость которого меньше 300 мг пара на 1 м2 за сутки. Такое полотно применяется во внутренних перегородках.
- Материал средней диффузии проводит от 300 до 1000 мг/м2 за сутки. Обычно применяют именно этот тип мембран, они хорошо зарекомендовали себя в зонах среднего и умеренного климата.
- Мембраны высокой диффузии или супердиффузионные имеют паропроницаемость более 1000 мг/м2/сутки. Они применяются в районах с большими перепадами влажности, суровым климатом или при большой толщине утеплителя.
Для чего предназначена диффузионная мембрана
Диффузионные мембраны защищают утеплитель, внутренние элементы конструкции кровли, стен и всего дома от потерь тепла. Водоотталкивающие свойства материала создают хорошую защиту от внешних воздействий, а воздухонепроницаемость позволяет уменьшить потери тепла в зимний период.
Мембрана защищает утеплитель от негативного воздействия влаги, которая проникает извне под наружную облицовку. Высокая паропроницаемость быстро удаляет воду из-под кровельного пирога.
Мембрана проводит влагу только в одностороннем направлении — от утеплителя, затем она стекает или испаряется. Стороны мембраны маркируются разными цветами, с нанесением логотипа или рисунка на одной из сторон. Ошибочный монтаж материала противоположной стороной обязательно приведет к намоканию теплоизолятора и нарушению утепления дома.
Где используется диффузионная мембрана
Область применения мембран довольно широкая: они могут быть использованы для гидроизоляции крыш, у которых есть подкровельная вентиляция, для теплоизоляционной защиты стропил по всей их высоте, в вентиляционных навесных фасадах и в несущих конструкциях из дерева. При использовании такой мембраны, выполнять дополнительную пропитку дерева необязательно.
Также диффузионная мембрана используется в вентилируемых помещениях и скатных кровлях, не имеющих утепления, где кровельным материалом является:
- полимерная черепица;
- шифер;
- керамическая черепица;
ондулин; - металлочерепица.

Преимущества диффузионных мембран
- надежность и высокая прочность материала;
- стойкость к перепадам температуры. Даже сильный перегрев мембраны не приводит к образованию вредных веществ;
- химическая стойкость к кислотам и щелочам;
- безопасность для здоровья.
Особенности монтажа диффузионной мембраны
Важно смонтировать мембрану нужной стороной (в случае с Ондутис — логотипом наружу, а чистой стороной к утеплителю). При монтаже диффузионных мембран важно исключить попадание влаги на теплоизоляцию через стыки и неплотности. Также обязательно нужно изолировать места, где пересекаются проникающие элементы конструкции: вентиляционные короба, антенны, каминные и печные трубы. Там, где они пересекаются, в мембране делается надрез в форме трапеции.
Чтобы мембрана эффективно функционировала, стоит предусмотреть естественную изоляцию пространства под кровлей и обеспечить удаление водяного пара. За эту функцию отвечает вентилируемый зазор между пленкой и покрытием. Его толщина равна толщине обрешетки и контррейки (обычно 7-12 см). Чтобы обеспечить доступ свежему воздуху и удалить водяной пар, делают отверстия для вентиляции.
За эту функцию отвечает вентилируемый зазор между пленкой и покрытием. Его толщина равна толщине обрешетки и контррейки (обычно 7-12 см). Чтобы обеспечить доступ свежему воздуху и удалить водяной пар, делают отверстия для вентиляции.
При наружном утеплении стен диффузионные мембраны монтируются вплотную к внешней стороне теплоизоляции. Материалы крепятся на несущий каркас с помощью строительного степлера или оцинкованных гвоздей. Сверху монтируются контррейки для создания вентиляционного зазора между внутренней стороной облицовочного материала и поверхностью пленки.
Что такое супердиффузионная мембрана
Супердиффузионная мембрана — материал нового поколения. Она будет незаменимой в области с суровым климатом и высокой влажностью, предназначена для комплексной защиты теплоизоляции и отличается от обычной мембраны высокой паропроницаемостью (более 1000 мг/м2/24 часа).
Cупердиффузионные мембраны Ондутис SA115 и SA130 пропускают водяной пар, но задерживают воду и воздух, защищают утеплитель и внутренние элементы стен и кровли. Они сохраняют утеплитель, уменьшают потери тепла кровельного пирога и могут использоваться как временная кровля до монтажа окончательной крыши в течение двух месяцев.
Они сохраняют утеплитель, уменьшают потери тепла кровельного пирога и могут использоваться как временная кровля до монтажа окончательной крыши в течение двух месяцев.
6 голосов , пожалуйста, оцените статью:
ondutis.ru
Диффузионные мембраны: применение, монтаж
В монтаже строительных конструкций помимо базовых свойств и эксплуатационных качеств используемых материалов важную роль играют вспомогательные элементы. В их числе особое место занимают изоляторы, обеспечивающие целостность основной структуры. Существенна функция данного компонента в кровельных покрытиях. Благодаря изоляционной прослойке формируется барьер перед осадками и другими внешними воздействиями. На рынке представлены разные по характеристикам средства обеспечения такой защиты, но самым эффективным материалом являются гидроизоляционные диффузионные мембраны, которые закладываются непосредственно в конструкцию крыши. Но кровельными системами не ограничиваются сферы применения данного покрытия.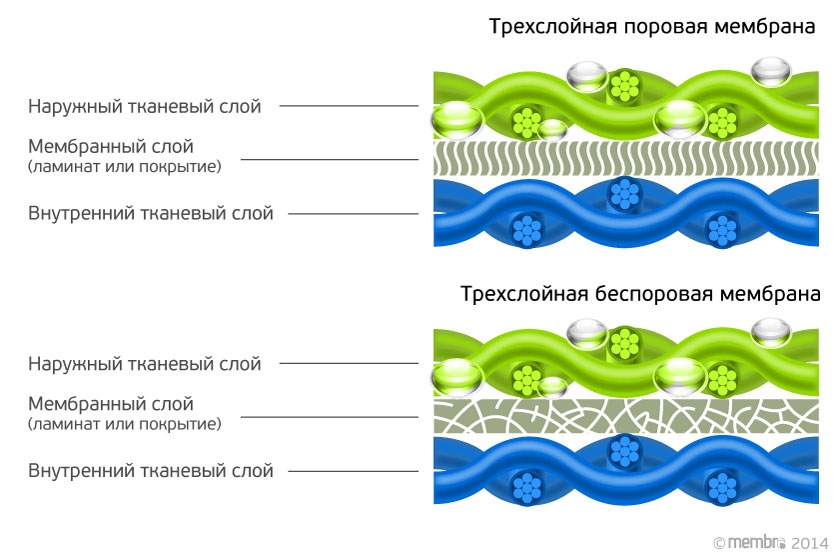 Например, изолятор также используют в укладке напольных и настенных материалов.
Например, изолятор также используют в укладке напольных и настенных материалов.
Общие сведения о диффузных изоляторах
Специально для сохранения технико-физических свойств настила кровли строители используют гидроизоляционные подкровельные пленки. Большинство из них реализуется в виде рулонов, которые внешне практически ничем не отличаются друг от друга. Оценивать же такие изоляторы можно по разным критериям, среди которых паропроницаемость. Собственно, это основное качество полимерных пленок, которое обуславливает возможность их структуры пропускать водяной пар. На языке специалистов этот параметр называется плотностью диффузионного потока. На практике использования паропроницаемость обуславливает интенсивность прохождения влаги через крышу в используемые помещения. Таким образом, диффузионные мембраны минимизируют возможность проникновения осадков и их производных под кровлю. Аналогичные функции выполняют практически все специализированные гидроизоляторы, но к мембранам относят и способность противостоять интенсивным воздействиям осадков в условиях порывистого ветра. Также если обычные изоляционные прослойки формируют лишь базовую защиту от дождевых капель и снега, то диффузионная пленка становится эффективной преградой перед водяным паром.
Также если обычные изоляционные прослойки формируют лишь базовую защиту от дождевых капель и снега, то диффузионная пленка становится эффективной преградой перед водяным паром.
Принцип действия мембраны
Распространенной проблемой многих гидроизоляторов является скопление влаги в одном месте – как правило, перед непроницаемым слоем. Грамотный монтаж предусматривает специальные каналы отвода жидкости, но и этот способ не избавляет от всех проблем эксплуатации. По другому принципу работают диффузионные мембраны, включенные в состав кровельных «пирогов». Он действует не пассивно, а активно, то есть не обеспечивают стационарный барьер, а за счет специального слоя проводит влагу от утеплителя к месту будущего стекания или выветривания. Поэтому очень важно правильной стороной располагать материал в рабочей зоне.
Сама структура функционального полотна таких изоляторов напоминает капиллярный насос, который формируется двумя прослойками. Ворсистая ткань выполняет функцию сборщика влаги и передает ее к участкам с перфорацией. Далее по микроскопическим отверстиям капли с паром просачиваются на внешнюю сторону полотна. В такой схеме влагоотвода есть риск контакта жидкости с металлическими элементами кровельной конструкции. Чтобы предотвратить процессы коррозии, используется объемная диффузионная мембрана на полипропиленовой нетканой основе. Также сберечь восприимчивые к влажности материалы помогут пленки с повышенной пропускной способностью – они быстрее обеспечивают вывод влаги, сокращая шансы возникновения разрушительных процессов.
Далее по микроскопическим отверстиям капли с паром просачиваются на внешнюю сторону полотна. В такой схеме влагоотвода есть риск контакта жидкости с металлическими элементами кровельной конструкции. Чтобы предотвратить процессы коррозии, используется объемная диффузионная мембрана на полипропиленовой нетканой основе. Также сберечь восприимчивые к влажности материалы помогут пленки с повышенной пропускной способностью – они быстрее обеспечивают вывод влаги, сокращая шансы возникновения разрушительных процессов.
Разновидности мембран
Различаются покрытия этого типа по нескольким критериям, но главным считается степень паропроницаемости. В частности, выделяют низко- и высокопаропроницаемые изоляторы. В первую категорию входят диффузионные мембраны, которые состоят из 2-3 слоев с армирующей сеткой. Такой материал обладает средними способностями к проведению водяного пара, но как универсальное средство защиты от осадков может стать оптимальным решением. Высокопаропроницаемые материалы демонстрируют более высокий уровень пропускной способности, из-за чего их также называют супердиффузионными пленками. Опять же, высокая скорость проведения влаги не позволяет ей конденсироваться, что способствует предохранению утепляющего слоя от разрушения. По этой причине строители упрощают изоляционные прослойки, избавляя их от вентиляционных зазоров.
Опять же, высокая скорость проведения влаги не позволяет ей конденсироваться, что способствует предохранению утепляющего слоя от разрушения. По этой причине строители упрощают изоляционные прослойки, избавляя их от вентиляционных зазоров.
Также в некоторых случаях используются специальные виды паропроницаемых пленок, среди которых и упомянутые объемные мембраны. Этот материал обычно применяют для обеспечения изоляции фальцевых и металлических покрытий. Монтируют объемные разделительные диффузионные мембраны с обеспечением верхнего вентиляционного зазора на кровлях, изготовленных из алюминия, стали, титана и других металлов.
Характеристики диффузных мембран
Основным эксплуатационным показателем таких материалов является паронепроницаемость. Но при выборе также учитываются и другие характеристики – например, грамматура, водонепроницаемость и температурная стойкость. Что касается грамматуры, то она обозначает массу пленки, которая увеличивается пропорционально толщине материала. Также по мере увеличения веса повышается и механическая прочность изоляции. На рынке можно найти пленки, грамматура которых варьируется от 60 до 270 г/м2. Также имеет значение и устойчивость покрытия к водяному давлению. Этой характеристикой обозначается водонепроницаемость, которой обладает диффузионная мембрана в конкретном исполнении. Существуют разные методики оценки этого параметра, но самой простой считается система, предусматривающая маркировку по классам W1-W3. Так, принадлежность мембраны к категории W1 указывает на то, что материал в течение 2 часов сможет выдерживать давление водяного столба, высота которого составляет 20 см.
Также по мере увеличения веса повышается и механическая прочность изоляции. На рынке можно найти пленки, грамматура которых варьируется от 60 до 270 г/м2. Также имеет значение и устойчивость покрытия к водяному давлению. Этой характеристикой обозначается водонепроницаемость, которой обладает диффузионная мембрана в конкретном исполнении. Существуют разные методики оценки этого параметра, но самой простой считается система, предусматривающая маркировку по классам W1-W3. Так, принадлежность мембраны к категории W1 указывает на то, что материал в течение 2 часов сможет выдерживать давление водяного столба, высота которого составляет 20 см.
Кроме осадков и влажности, вред гидроизоляционным пленкам способна нанести теплая солнечная погода. В стандартном исполнении такие материалы выдерживают пиковое температурное воздействие в диапазоне 80-100 °C. Что касается стойкости к ультрафиолету, то, например, диффузионная мембрана «Технониколь» в базовой версии может находиться под прямыми лучами солнца 3 месяца. Для усиленных модификаций этот временной промежуток увеличивается до 4 месяцев.
Для усиленных модификаций этот временной промежуток увеличивается до 4 месяцев.
Применение в кровельной конструкции
В составе кровли изоляционная мембрана обеспечивает защиту утеплителя и внутренних частей конструкции. Прослойка выполняет несколько функций одновременно – кроме предохранения от влаги и пара она также ограждает элементы крыши от выветривания. Неправильно относить задачи данного материала к изоляции помещений здания. Основные функции мембраны все же распространяются на защиту утепляющего слоя, который, в свою очередь, регулирует влажностный, а в некоторых случаях и температурный режимы. В ходе проектирования схемы устройства мембраны необходимо предотвратить возможности стекания воды с пленочной поверхности на теплоизолятор по местам стыков. Как правило, подкровельная диффузионная мембрана соседствует с каналами вентиляции, каминными и печными трубами, стойками антенн и другими сторонними компонентами, пересечения с которыми необходимо изолировать.
Применение мембран в устройстве напольных покрытий
Обеспечение надежной изоляции требуется и в самих помещениях. Так, напольные поверхности сохраняют декоративные и технические качества только в условиях оптимального температурно-влажностного режима. Конечно, есть материалы, которые не чувствительны к тем же воздействиям влаги, но древесные поверхности к таким не относятся. Обычно применение диффузионных мембран в напольных покрытиях ограничивается созданием вспомогательной подложки для утеплителя. Впрочем, может иметь значения этаж, на котором выполняются работы. Самым требовательным является нижний уровень в частном доме, особенно если он связывает жилые помещения с подвалом.
Так, напольные поверхности сохраняют декоративные и технические качества только в условиях оптимального температурно-влажностного режима. Конечно, есть материалы, которые не чувствительны к тем же воздействиям влаги, но древесные поверхности к таким не относятся. Обычно применение диффузионных мембран в напольных покрытиях ограничивается созданием вспомогательной подложки для утеплителя. Впрочем, может иметь значения этаж, на котором выполняются работы. Самым требовательным является нижний уровень в частном доме, особенно если он связывает жилые помещения с подвалом.
Применение в облицовке стен и фасадов
С помощью диффузионной пленки также обеспечивают защиту от выветривания и влаги теплоизоляционных материалов, находящихся в составе вентилируемых фасадов. Мембраны размещаются с внешней стороны, обеспечивая, таким образом, возможность прохождения излишков влаги и пара через вентилируемый зазор в облицовке. Установка производится непосредственно на фасаде дома вместе с теплоизоляционными панелями. Кстати, если в конструкции используется металлический несущий каркас, то оптимально подойдет объемная диффузионная мембрана, которая создаст естественный сток влаги без риска развития коррозии. Если планируется использовать в отделке сайдинг или вагонку, то пленку нужно фиксировать вплотную к наружной стороне изоляции под декоративной облицовкой. Это один из простейших способов применения диффузных изоляторов, так как он требует минимальных усилий в плане монтажа – достаточно воспользоваться строительным степлером или оцинкованными гвоздиками и закрепить материал к несущей конструкции.
Кстати, если в конструкции используется металлический несущий каркас, то оптимально подойдет объемная диффузионная мембрана, которая создаст естественный сток влаги без риска развития коррозии. Если планируется использовать в отделке сайдинг или вагонку, то пленку нужно фиксировать вплотную к наружной стороне изоляции под декоративной облицовкой. Это один из простейших способов применения диффузных изоляторов, так как он требует минимальных усилий в плане монтажа – достаточно воспользоваться строительным степлером или оцинкованными гвоздиками и закрепить материал к несущей конструкции.
Монтаж диффузионной мембраны
При наружном способе размещения материала желательно выполнять операцию в сухую погоду и вместе с фиксацией теплоизолятора. Мембрану следует раскатать по всей рабочей поверхности и закрепить ее гвоздиками или скобками. Классическая установка на крыше предполагает укладку горизонтальными полосками с нахлестом порядка 15 см. При этом вертикальные стыки окончаний следует размещать на стропилах. Все точки и линии промежутков между отдельными кусками материала фиксируются с помощью бутилкаучуковой или акриловой ленты. Вентиляционный зазор не требуется, если диффузионные мембраны планируется устанавливать на крыше, но при укладке изолятора на фасадных поверхностях его лучше предусмотреть. В зимнее время нагрузка на утепляющий материал значительно повышается, поэтому рекомендуется осенью выполнять ревизию кровли на предмет плотности прилегания мембраны к изолятору. Если у стропил толщина превышает аналогичный показатель теплоизолятора, то паронепроницаемый материал следует прижать к их боковой стороне рейками с типоразмером 2х3 см. Также для выполнения этой задачи можно использовать строительные скобы. В ходе работы следует ориентироваться на то, чтобы нижняя кромка мембраны обеспечивала беспрепятственное удаление влаги в специальный желоб водоотвода.
Все точки и линии промежутков между отдельными кусками материала фиксируются с помощью бутилкаучуковой или акриловой ленты. Вентиляционный зазор не требуется, если диффузионные мембраны планируется устанавливать на крыше, но при укладке изолятора на фасадных поверхностях его лучше предусмотреть. В зимнее время нагрузка на утепляющий материал значительно повышается, поэтому рекомендуется осенью выполнять ревизию кровли на предмет плотности прилегания мембраны к изолятору. Если у стропил толщина превышает аналогичный показатель теплоизолятора, то паронепроницаемый материал следует прижать к их боковой стороне рейками с типоразмером 2х3 см. Также для выполнения этой задачи можно использовать строительные скобы. В ходе работы следует ориентироваться на то, чтобы нижняя кромка мембраны обеспечивала беспрепятственное удаление влаги в специальный желоб водоотвода.
Производители
Нетканый паронепроницаемый материал представляют на рынке разные изготовители, среди которых немало и отечественных. В частности, диффузионная мембрана «Технониколь» позволяет устраивать гидроизоляцию, которая одновременно защищает конструкции от контактов с влагой и дает поверхностям «дышать». Материал этой линейки представлен в нескольких версиях, в том числе и с усиливающими волокнами. Также у пользователей завоевала доверие супердиффузионная пленка «Технониколь», которая выполнена из трехслойного микропористого материала. Отличием данного изолятора является двухстороннее применение нетканого полипропилена, выполняющего защитную функцию для основной прослойки. Такую мембрану целесообразно использовать именно в кровельных покрытиях, которые требуют не только качественного выполнения изоляционных задач, но и сохранения прочностных свойств конструкции.
В частности, диффузионная мембрана «Технониколь» позволяет устраивать гидроизоляцию, которая одновременно защищает конструкции от контактов с влагой и дает поверхностям «дышать». Материал этой линейки представлен в нескольких версиях, в том числе и с усиливающими волокнами. Также у пользователей завоевала доверие супердиффузионная пленка «Технониколь», которая выполнена из трехслойного микропористого материала. Отличием данного изолятора является двухстороннее применение нетканого полипропилена, выполняющего защитную функцию для основной прослойки. Такую мембрану целесообразно использовать именно в кровельных покрытиях, которые требуют не только качественного выполнения изоляционных задач, но и сохранения прочностных свойств конструкции.
Довольно известным изготовителем паронепроницаемых гидроизоляторов является и компания Du Pont, специализирующаяся как раз на защите стеновых и кровельных систем посредством воздушной и водной фильтрации. Под этим брендом выходит диффузионная мембрана Tyvek, представляющая собой многофункциональную гидроветрозащитную пленку.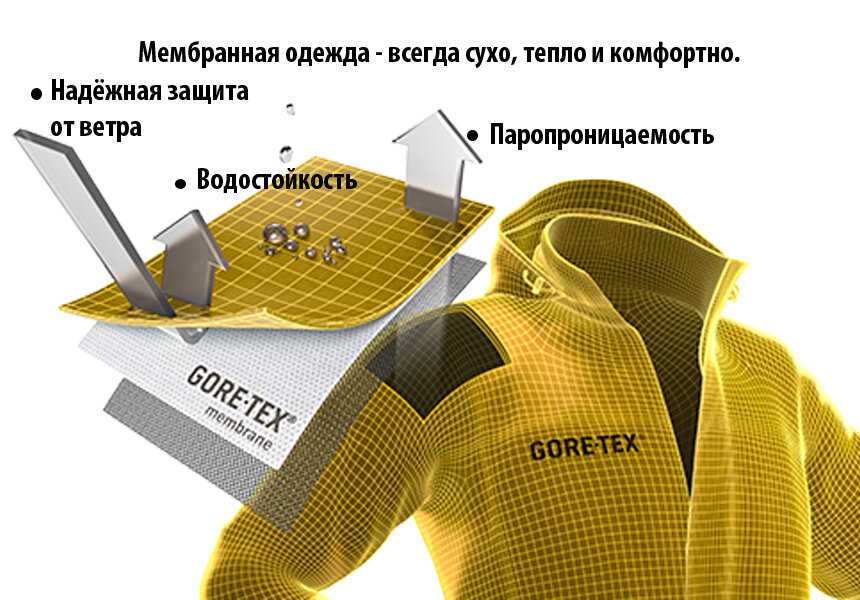 Материал имеет сетчатую структуру, образованную полимерными волокнами. В отличие от конкурирующих изоляторов покрытия Tyvek отличаются экологической чистотой. То есть их можно использовать и в наружной отделке, и в структуре внутренних облицовочных покрытий. В обоих случаях включение диффузионной пленки этой марки будет обеспечивать сухость и долговечность отделки.
Материал имеет сетчатую структуру, образованную полимерными волокнами. В отличие от конкурирующих изоляторов покрытия Tyvek отличаются экологической чистотой. То есть их можно использовать и в наружной отделке, и в структуре внутренних облицовочных покрытий. В обоих случаях включение диффузионной пленки этой марки будет обеспечивать сухость и долговечность отделки.
Заключение
Традиционные способы устройства изоляционных прослоек обычно не требуют соблюдения особых условий монтажа. Достаточно подобрать лист или плиту соответствующих размеров и зафиксировать материал на рабочем участке. По такому принципу обычно монтируется и утепляющий материал, и гидроизоляция. Диффузионная мембрана, в свою очередь, гарантирует более высокий эксплуатационный эффект, чем обычные изоляторы. Но для его достижения важно соблюдать правила расположения и крепления паронепроницаемых пленок относительно утепляющих слоев. Кровельные конструкции в этом отношении являются наиболее сложными, так как выводящий влагу материал может контактировать с другими компонентами покрытия и системами крыши. Также изначально следует предусмотреть коммуникации для водоотвода, с которыми будет взаимодействовать диффузионный материал. С точки зрения установки, несколько проще выглядит монтаж изоляторов на стенах и фасадных конструкциях. В таких случаях все-таки на первое место выходит задача сохранения вентиляционных свойств покрытия, которые и реализуются благодаря включению паронепроницаемой мембраны в состав утепляющей прослойки.
Также изначально следует предусмотреть коммуникации для водоотвода, с которыми будет взаимодействовать диффузионный материал. С точки зрения установки, несколько проще выглядит монтаж изоляторов на стенах и фасадных конструкциях. В таких случаях все-таки на первое место выходит задача сохранения вентиляционных свойств покрытия, которые и реализуются благодаря включению паронепроницаемой мембраны в состав утепляющей прослойки.
fb.ru
Что такое диффузионная мембрана, плюсы и минусы
>
Вступление
Важнейшим слоем в «пироге» многослойной конструкции кровли является слой гидроизоляции. Лучшим материалом для гидроизоляции кровли является диффузионная мембрана.
Диффузионная мембрана это
Диффузионная мембрана – материал, используемый для гидроизоляции кровли, предохранения от попадания влаги в утеплитель и производства других работ в строительстве. Данный продукт новых технологий появился сравнительно недавно.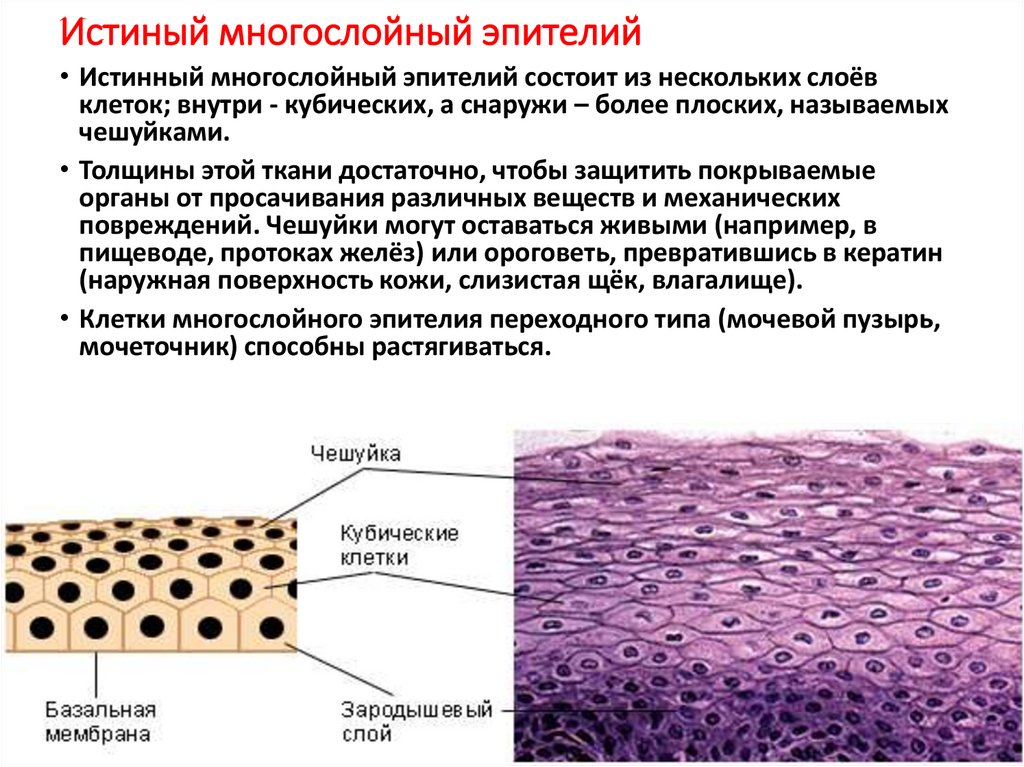 От обыкновенных пленок (использовавшихся ранее с той же целью) отличается своей способностью к однонаправленному пропусканию влаги. При этом не пропускает ни воду, ни воздух, работая одновременно и тепло и гидроизолятором.
От обыкновенных пленок (использовавшихся ранее с той же целью) отличается своей способностью к однонаправленному пропусканию влаги. При этом не пропускает ни воду, ни воздух, работая одновременно и тепло и гидроизолятором.
Устройство диффузионной мембраны
Данный материал предлагается в виде рулонов (для удобства использования). Многослойное полотно, представляющее собой капиллярный насос (влага перемещается с ворсистой поверхности к гладкой). Мембраны Ондутис (продукция компании Ондулин) 3-слойные – снаружи нетканый полипропилен, внутри стойкий к растяжению усилитель. Благодаря такой конструкции, мембрана этой марки имеет не только высокие характеристики гидрофобности и паропроницаемости, но и отличную механическую прочность. Слои мембраны соединяются в пакет посредством высокотехнологичной ультразвуковой сварки. Материал экологически безопасен и устойчив к воздействию микроорганизмов.
Свойства диффузионной мембраны
Важнейшим свойством диффузионной мембраны является её паропроницаемость. Измеряется она в миллиграммах пара, проходящих через квадратный метр поверхности за 24 часа. В зависимости от ее величины различают мембраны:
Измеряется она в миллиграммах пара, проходящих через квадратный метр поверхности за 24 часа. В зависимости от ее величины различают мембраны:
- Мало диффузионные. Менее 300 мг/м2/сутки. Используются только внутри помещений.
- Средне диффузионные. 300-1000 мг/м2/сутки. Наиболее массовые в использовании. Отлично работают в зонах с умеренным климатом.
- Высоко диффузионные (супердиффузионные). Свыше 1000 мг/м2/сутки. Используются при строительстве в сложных климатических условиях (или при использовании толстого утеплителя), в местах с резкой сменой влажности.
С какой целью разработана диффузионная мембрана
Материал был разработан для защиты утеплителя от промокания (и как следствие, потери теплоизоляционных свойств), сохранения в рабочем состоянии стропил, и других внутренних элементов кровли, защиты стен и сохранения тепла в доме, в целом. Материал воздухопроницаем, благодаря чему дом дышит, и не нуждается в дополнительном проветривании. В то же время не пропускает воду, защищая жилище от непогоды. Благодаря высокой паропроницаемости происходит быстрое удаление водного конденсата из-под кровли или наружной облицовки стен.
В то же время не пропускает воду, защищая жилище от непогоды. Благодаря высокой паропроницаемости происходит быстрое удаление водного конденсата из-под кровли или наружной облицовки стен.
Мембрана обладает свойством односторонней проницаемости. Вода забирается от утеплителя, затем по другой стороне стекает или испаряется. С учетом этого при производстве стороны красят в разный цвет. А на одну из сторон наносятся надписи, пиктограммы, фирменные изображения. Ошибка в выборе правильной ориентации материала при монтаже приведет к тому, что теплоизоляция намокнет, и ее свойства резко ухудшатся.
Где используется диффузионная мембрана
Мембраны массово применяются в строительстве зданий и сооружений для гидроизоляции крыш (при наличии подкровельной вентиляции), теплоизоляции и влагозащиты стропил. Защищают несущие деревянные конструкции. Используют в вентиляцонных навесных фасадах. При использовании такого материала в деревянном домостроении НЕ требуется дополнительная пропитка древесины антисептиком.
Кроме того, диффузионная мембрана применяется при обустройстве не утепленной кровли из:
- полимерной или керамочерепицы;
- металлочерепицы;
- шифера;
- ондулина.
Сильные стороны диффузионных мембран
К основным преимуществам данной продукции можно отнести:
class=”eliadunit”>
- высокую механическая прочность;
- температурную выносливость (хорошо выносит перепады температуры), при нагреве – не выделяет опасных элементов;
- устойчивость к химически активным реагентам;
- экологичность.
Тонкости укладки диффузионной мембраны
Укладка полотна обязательно проводится с учетом направления работы мембраны. Продукция Ондутис кладется логотипом наружу, поверхностью без надписей – к утеплителю. Все стыки и неплотные прилегания должны быть надежно заизолированы, во избежание проникновения воды в утеплитель. Необходимо тщательно изолировать места выходов труб, антенн и других проникающих конструкций. В мембране под них производится трапецеидальный надрез.
В мембране под них производится трапецеидальный надрез.
Для нормальной работы мембраны, надо заблаговременно обеспечить естественную вентиляцию под кровлей с целью отвода водяного пара, создать продуваемый промежуток между ней и покрытием. Размер зазора обычно составляет 70-120мм (сумма толщин брусков обрешетки и контррейки). Для доступа наружного воздуха предусматривают вентиляционные отверстия.
Если мембрана используются для утепления стен снаружи, ее монтируют прямо к утеплителю. Крепление мембраны производится в строительный каркас (сквозь теплоизоляцию) скобами или гвоздями. Поверх устанавливаются направляющие, к которым крепится облицовка. В итоге между облицовкой и пленкой создается необходимый вентиляционный зазор.
Супердиффузионная мембрана – что это и зачем нужна
Инновационный продукт, созданный для условий с экстремальными температурами и влажностью, назначение, которого – эффективная защита теплоизоляции с высокими показателями паропроницаемости.
Новые продукты – супермембраны Ондутис SA115 и SA130 еще лучше пропускают пар, но обеспечивают надежную преграду воде и воздуху. Надежно оберегают теплоизоляцию и нелюбящие влажность конструктивные элементы зданий. Помогая сохранять свойства теплозащиты, они снижают затраты на поддержание тепла в доме.
©DomiTy.ru
Статьи по теме
>
domity.ru
Диффузионные мембраны: виды, характеристики, применение
Статья расскажет о современном материале для парогидроизоляции — диффузионных мембранах. Вы узнаете о том, какие бывают гидробарьеры и в каких случаях применяется каждый из них. Также в статье есть таблица с анализом рыночных цен на диффузионные мембраны.
Гидроизоляционный рулонный материал, применяемый для наружных работ, должен иметь одно свойство — одностороннюю проводимость влаги. Это необходимо для того, чтобы удалять из утеплителя пары, образованные в результате перепадов температуры и повышения влажности. Единственным материалом, способным выполнять эту работы является, мембрана.
Единственным материалом, способным выполнять эту работы является, мембрана.
Как работает мембрана
Возможные варианты названия — проводящая, полупроводящая, диффузионная, супердиффузионная и пароизоляцоинная мембрана (не плёнка!) отражают суть работы этого слоя. Мембрана проводит влагу в одну сторону — от утеплителя — и концентрирует её на обратной стороне для последующего стекания или выветривания. Поэтому стороны полотна мембраны маркированы однозначным образом — рисунок, логотип, разный цвет. Укладка этого материала обратной стороной (по ошибке) приведёт к тому, что утеплитель размокнет за считанные недели.
Само полотно представляет собой капиллярный насос, образованный двумя слоями материала:
- Ворсистый слой. Собирает влагу и передаёт к перфорации. Через микроскопические отверстия влага просачивается на обратную сторону полотна.
- Гладкий полиэтиленовый слой с перфорацией. На нём собирается отведённая от утеплителя влага.
От плотности перфорации (количества отверстий на единицу площади) напрямую зависит «пропускающая способность» полотна. Чем их больше, тем выше способность мембраны отводить воду.
Чем их больше, тем выше способность мембраны отводить воду.
Паропроницаемость — главное свойство мембраны
По этому признаку все мембраны делятся на три категории:
- Малой диффузии — проводят менее 300 мг на 1 м2 в течение 24 часов. Такое полотно подходит для сухих помещений и внутренних перегородок.
- Средней диффузии — от 300 до 1000 мг/м2/24 ч. Этот вид мембраны подходит в большинстве случаев — в среднем и умеренном климате.
- Высокой диффузии (супердиффузионные) — более 1000 мг/м2/24 ч. Используется при значительной толщине утеплителя, в районах с суровым климатом и перепадами влажности.
Этот интересный эффект успешно используют также в растениеводстве — для посадок в теплицах, которые не любят влажного воздуха. Мембрана осушает закрытую атмосферу такого «парника».
Монтаж мембраны
Пароизоляция, уложенная между кровельным материалом или вентилируемым фасадом и конструктивом стены или кровли, условно называется гидроизоляцией или гидробарьером. Этот термин условный, т. к. отражает не свойства материала, а его назначение. В данном случае мембрана действительно не пропускает влагу, но назвать её полноценной гидроизоляцией не получается.
Этот термин условный, т. к. отражает не свойства материала, а его назначение. В данном случае мембрана действительно не пропускает влагу, но назвать её полноценной гидроизоляцией не получается.
Как было сказано выше, производитель делает всё для того, чтобы не произошло ошибки при монтаже, и мембрана была установлена правильно. Но это далеко не все требования к ней. Укладка полотна диффузионной мембраны требует соблюдения следующих правил:
- Полотно раскатывается на уложенный утеплитель ворсистой стороной к нему и при этом плотно прилегает.
- Полотно прижимается к стропилам или стойкам прижимным брусом или контробрешёткой.
- Между гидробарьером и отделочным материалом кровли или стены должен быть продух 15–50 мм.
- Образованный прижимными брусками канал должен быть свободным от софита до конька (на кровле) или стартовой рейки или софита (по стене).
- Если используется контробрешётка, в ней предусматриваются разрывы для сообщения воздушных каналов между собой.
- Полотна гидробарьера необходимо надёжно склеивать между собой, соблюдая нахлёст 50–100 мм. Крайние примыкания к обрешётке также проклеиваются.
Правильно смонтированная мембрана прослужит очень долго — от 50 лет. Отсутствие этого слоя в кровельном пироге повлечёт за собой нарушение работы утеплителя.
Специальные мембраны
К этой категории относятся высокотехнологичные материалы, которые комбинируют в себе свойства паробарьера и утеплителя.
Металлизированная мембрана. Разработка фирм Tyvek и DuPont. Алюминий, напылённый на каждое волокно в отдельности, обеспечивает частичное отражение тепла при сохранении свойств диффузии.
Фольгированная мембрана. Особенно актуальна для жарких районов. Фольга отражает инфракрасное тепло обратно в атмосферу. В результате крыша греется в разы меньше. Зимой такой барьер поддерживает температуру в вентиляционном продухе для испарения конденсата.
Цены на диффузионные мембраны
| Название | Производитель | Вес 1 м2, г | Паропроницаемость, мг/м2/24 ч | Площадь рулона, м2 | Цена рулона, у. е. е. | Цена 1 м2, у. е. |
| BRANE SM | Россия | 75 | 1000 | 70 | 26,5 | 0,37 |
| Ондутис SA115 | Россия | 115 | 1100 | 75 | 30 | 0,4 |
| РуфИзол SD | Россия | 100 | 1250 | 70 | 35,5 | 0,5 |
| Строизол SD 130 | Россия | 130 | 1500 | 70 | 38,5 | 0,53 |
| CHESCO SD | Россия | 115 | 1200 | 60 | 37 | 0,61 |
| Fakro Eurotop N35 | США | 135 | 1900 | 75 | 58,5 | 0,78 |
| DELTA VENT N | Германия | 120 | 1750 | 75 | 70 | 0,93 |
| Tyvek Solid Silver (металлизированная) | Лихтенштейн | 82 | 1300 | 75 | 110 | 1,47 |
| Изовек Т (фольгированная) | Россия | 150 | 1000 | 60 | 58,5 | 0,97 |
Первая мысль, которая приходит в голову при попытке сэкономить на гидробарьере — заменить диффузионную мембрану обычным полиэтиленом. Такой вариант будет выполнять функцию на 50% (останавливать атмосферную влагу) и на 150% будет портить всё дело. Невыпущенный из утеплителя пар накопится в нём и размочит вату. А вата, как известно, восстановлению не подлежит. Поэтому применяйте диффузионные мембраны, чтобы кровля и стены были всегда сухими и теплыми.
Такой вариант будет выполнять функцию на 50% (останавливать атмосферную влагу) и на 150% будет портить всё дело. Невыпущенный из утеплителя пар накопится в нём и размочит вату. А вата, как известно, восстановлению не подлежит. Поэтому применяйте диффузионные мембраны, чтобы кровля и стены были всегда сухими и теплыми.
Виталий Долбинов, рмнт.ру
rmnt.mirtesen.ru
Какую выбрать супердиффузионную мембрану?
Как известно, любая кровельная конструкция представляет собой многослойную структуру, в которой слой утеплителя должен быть обязательно защищен от проникновения влаги паро- и гидроизоляционными пленками. Какими характеристиками должны обладать пленки, предназначенные для защиты утепляющего слоя, учитывая, что даже при абсолютной целостности крыши влага все равно попадает в подкровельное пространство? Как правило, влага проникает в пространство под кровлей после дождя или тумана, она может конденсироваться в результате наличия разности наружной и внутренней температуры воздуха, а также просачиваться в ходе диффузионных процессов и конвекции из нижерасположенных помещений. Наличие влаги отрицательно сказывается на теплосберегающих характеристиках утеплителя. Как показывают результаты исследований, увлажнение теплоизоляционного слоя всего лишь на 1% способно снизить характеристики эффективности слоя изоляции на 34%!
Наличие влаги отрицательно сказывается на теплосберегающих характеристиках утеплителя. Как показывают результаты исследований, увлажнение теплоизоляционного слоя всего лишь на 1% способно снизить характеристики эффективности слоя изоляции на 34%!
Что такое супердиффузионные мембраны?
Диффузионная мембрана – это специальный материал, имеющий двух-, трех- или даже четырехслойную структуру, основу которого составляет нетканый холст. Диффузионные мембраны применяют для защиты утепляющего слоя от проникновения в его толщу испарений. Также, диффузионные мембраны являются превосходной защитой от воды и ветра. При создании крыши, в полном объеме соответствующей всем современным требованиям, каждый застройщик обязательно столкнется с таким понятием, как «кровельный пирог». Для того чтобы крыша выполняла все возложенные на нее функции в течение всего срока эксплуатации, кроме основного кровельного покрытия, необходимо использовать некоторые дополнительные материалы, к числу которых относятся супердиффузионные мембраны. Супердиффузионные мембраны можно использовать при создании кровельного пирога в любой климатической зоне нашей страны. Роль этого дополнительного слоя чрезвычайно важна, так именно его присутствие позволяет снизить силу неблагоприятных воздействий, вызванных экстремальными погодными условиями, а также нивелировать недочеты и ошибки, возникшие в ходе неправильного монтажа кровли.
Супердиффузионные мембраны можно использовать при создании кровельного пирога в любой климатической зоне нашей страны. Роль этого дополнительного слоя чрезвычайно важна, так именно его присутствие позволяет снизить силу неблагоприятных воздействий, вызванных экстремальными погодными условиями, а также нивелировать недочеты и ошибки, возникшие в ходе неправильного монтажа кровли.
Предназначение диффузионных мембран
Основное предназначение супердиффузионных мембран для кровли является обеспечение защиты от проникновения внутренней и наружной влаги внутрь теплоизоляционного слоя. Источниками этой влаги могут быть внутренние испарения и атмосферные осадки. Кроме этого, расположенная в кровельном покрытии диффузионная мембрана обеспечивает эффективные условия отвода уже накопившейся в силу тех или иных причин влаги. Супердиффузионную мембрану можно с полной уверенностью назвать одной из важнейших составляющих теплоизоляционного контура, так как она косвенным образом способствует снижению потерь тепловой энергии. Бережливый хозяин собственного дома, знающий толк в экономии, никогда не будет раздумывать о необходимости или отсутствии таковой при принятии решения о покупке и последующей установке диффузионной мембраны. Тем более, что стоимость этого материала на современном рынке строительных материалом можно с уверенностью назвать чисто символической.
Бережливый хозяин собственного дома, знающий толк в экономии, никогда не будет раздумывать о необходимости или отсутствии таковой при принятии решения о покупке и последующей установке диффузионной мембраны. Тем более, что стоимость этого материала на современном рынке строительных материалом можно с уверенностью назвать чисто символической.
Преимущества использования супердиффузионных мембран
Хозяин частного дома, решивший использовать в конструкции кровельного пирога супердиффузионные мембраны, в сравнении с домовладельцами, использующими традиционные технологии, получит ряд неоспоримых преимуществ, среди которых основными можно назвать следующие:
- Использование супердиффузионных мембран позволяет одной пленке заменить две, такие как гидро- и ветрозащита. Наличие мембраны допускает возведение конструкции без наличия вентиляционного зазора.
- Укладка супердиффузионных мембран разрешается непосредственно на поверхность любого покрытия, что позволяет укладывать теплоизоляцию более толстым слоем, в сравнении с традиционными технологиями.
 Как результат, владелец дома получает усиленную теплоизоляцию.
Как результат, владелец дома получает усиленную теплоизоляцию. - Использование супердиффузионных мембран позволяет продлить срок эксплуатации утепляющего материала и деревянных конструкций кровли. При этом, деревянные элементы крыши могут быть установлены без предварительной обработки специальными химическими составами.
- Применение супердиффузионных мембран в ходе создания кровельного пирога значительно сокращает время проведения монтажных работ и связанных с ними затрат.
Разновидности супердиффузионных мембран
В свободной продаже супердиффузионные мембраны можно встретить в виде рулонов. Рулоны имеют незначительную массу, благодаря чему проведение работ, связанных с благоустройством кровли, проходит достаточно просто. Диффузионные мембраны обладают следующими основными характеристиками: количество слоев, степень проницаемости пара, плотность материала, водопроницаемость. Необходимый для конкретных условий материал необходимо выбирать на основе числовых значений этих показателей. В настоящее время на рынке присутствуют следующие разновидности супердиффузионных мембран:1. Difoil-S. Диффузионная пленка этого типа имеет трехслойную структуру. Материал характеризуется высокой степенью паропроницаемости, он способен достаточно быстро отводить из помещения водяной пар. Чаще всего эту пленку укладывают непосредственно на теплоизолирующий материал или на поверхность настила, при этом необходимость создания вентиляционного зазора отсутствует. Максимально успешно данная супердиффузионная пленка может быть использована для мансардных крыш, она превосходно сочетается с обыми типами кровельных материалов, присутствующих в настоящее время на строительном рынке. Difoil-S способен обеспечить эффективную защиту крыши и стен от влаги и ветровых нагрузок. Чаще всего, этот материал используют в ходе утепления наружных стен. В вентилируемых кровельных конструкциях данную диффузионную пленку укладывают непосредственно на поверхность теплоизоляционного материала, что позволяет создать дополнительную защиту от проникновения атмосферной влаги.
Необходимый для конкретных условий материал необходимо выбирать на основе числовых значений этих показателей. В настоящее время на рынке присутствуют следующие разновидности супердиффузионных мембран:1. Difoil-S. Диффузионная пленка этого типа имеет трехслойную структуру. Материал характеризуется высокой степенью паропроницаемости, он способен достаточно быстро отводить из помещения водяной пар. Чаще всего эту пленку укладывают непосредственно на теплоизолирующий материал или на поверхность настила, при этом необходимость создания вентиляционного зазора отсутствует. Максимально успешно данная супердиффузионная пленка может быть использована для мансардных крыш, она превосходно сочетается с обыми типами кровельных материалов, присутствующих в настоящее время на строительном рынке. Difoil-S способен обеспечить эффективную защиту крыши и стен от влаги и ветровых нагрузок. Чаще всего, этот материал используют в ходе утепления наружных стен. В вентилируемых кровельных конструкциях данную диффузионную пленку укладывают непосредственно на поверхность теплоизоляционного материала, что позволяет создать дополнительную защиту от проникновения атмосферной влаги. Difoil-S превосходно сочетается с любыми типами изолирующих материалов. Монтируется данная мембрана с небольшим провисанием или сверху слоя теплоизоляции стороной, на которой имеются надписи, в направлении крыши.
Difoil-S превосходно сочетается с любыми типами изолирующих материалов. Монтируется данная мембрана с небольшим провисанием или сверху слоя теплоизоляции стороной, на которой имеются надписи, в направлении крыши.
2. MASTERMAX 3 ECO – супердиффузионная мембрана для подкровельного применения, характеризующаяся высоким качеством и превосходными эксплуатационными характеристиками. Этот материал используют в качестве защитного покрытия кровельной конструкции. Его использование позволяет создать надежную защиту утеплителя от влаги, ветра и пыли. Также, материал способен отводить от поверхности теплоизолятора испарения. В виду того, что диффузионная мембрана изготовлена на основе полипропиленового волокна, она обладает высокой паропроницаемостью и увеличенной способностью пропускания водяных паров из пространства под кровлей. Благодаря ниличию подробных характеристик, MASTERMAX называют мембранами новейшего поколения. Мембраны MASTERMAX 3 ECO могут эксплуатироваться с любыми видами теплоизоляционных материалов. Для получения максимальной эффективности, мембрану необходимо устанавливать правильно, а именно, нанесенная на поверхность материала надпись MASTERMAX 3 ECO должна быть расположена в сторону кровли. 3. MASTERMAX 3 CLASSIC. Данный материал представляет собой трехслойную гидроизоляционную супердиффузионную мембрану. Он используется для создания второстепенной защиты от воздействия влаги и пыли. Пленку монтируют сверху теплоизоляционного слоя. 4. MASTERMAX 3 TOP. Данная супердиффузионная мембрана предназначена для защиты слоя утеплителя тех кровель, которые эксплуатируются в условиях повышенных ветровых и влажностных нагрузок. Эту пленку, также как и прочие ее аналоги, устанавливают поверх слоя утеплителя. 5. MASTERMAX 3 EXTRA представляет собой высокотехнологичную мембрану, которую также как и MASTERMAX 3 TOP монтируют в пределах кровельного пирога крыш, подверженных экстремальным влажностным и ветровым нагрузкам. Данная разновидность мембран является наиболее плотной и самой высококачественной пленкой среди представителей трехслойных мембран от производителя MASTERMAX.
Для получения максимальной эффективности, мембрану необходимо устанавливать правильно, а именно, нанесенная на поверхность материала надпись MASTERMAX 3 ECO должна быть расположена в сторону кровли. 3. MASTERMAX 3 CLASSIC. Данный материал представляет собой трехслойную гидроизоляционную супердиффузионную мембрану. Он используется для создания второстепенной защиты от воздействия влаги и пыли. Пленку монтируют сверху теплоизоляционного слоя. 4. MASTERMAX 3 TOP. Данная супердиффузионная мембрана предназначена для защиты слоя утеплителя тех кровель, которые эксплуатируются в условиях повышенных ветровых и влажностных нагрузок. Эту пленку, также как и прочие ее аналоги, устанавливают поверх слоя утеплителя. 5. MASTERMAX 3 EXTRA представляет собой высокотехнологичную мембрану, которую также как и MASTERMAX 3 TOP монтируют в пределах кровельного пирога крыш, подверженных экстремальным влажностным и ветровым нагрузкам. Данная разновидность мембран является наиболее плотной и самой высококачественной пленкой среди представителей трехслойных мембран от производителя MASTERMAX. Кроме своего основного предназначения, пленка MASTERMAX 3 EXTRA способна успешно противостоять воздействию прямых ультрафиолетовых лучей. Данную мембрану укладывают без вентиляционного зазора. 6. MASTERMAX METAL – это пленка, которая относится к категории четырехслойных супердиффузионных мембран. Этот паропроницаемый материал используют в конструкции кровельного пирога, в котором он выполняет роль второстепенной защиты от снега, дождя, тумана, ветра и пыли. MASTERMAX METAL укладывают под металлические листы жестяных крыш, лежащих на деревянной основе. Материал применяется для кровель, подверженных значительным тепловым нагрузкам. Применение MASTERMAX METAL предоставляет идеальные условия для организации эффективной вентиляции пространства под кровлей. Пленка способна обеспечить непрерывную циркуляцию потоков воздуха, в результате чего влажный воздух, конденсат и пар максимально быстро отводятся из чердачного помещения. Более того, использование этой супердиффузионной мембранной пленки позволяет создать между крышей и утеплителем определенное термическое пространство, благодаря которому в летнее время года уменьшается нагрев дома, в зимний период на поверхности крыши не будет появляться наледь.
Кроме своего основного предназначения, пленка MASTERMAX 3 EXTRA способна успешно противостоять воздействию прямых ультрафиолетовых лучей. Данную мембрану укладывают без вентиляционного зазора. 6. MASTERMAX METAL – это пленка, которая относится к категории четырехслойных супердиффузионных мембран. Этот паропроницаемый материал используют в конструкции кровельного пирога, в котором он выполняет роль второстепенной защиты от снега, дождя, тумана, ветра и пыли. MASTERMAX METAL укладывают под металлические листы жестяных крыш, лежащих на деревянной основе. Материал применяется для кровель, подверженных значительным тепловым нагрузкам. Применение MASTERMAX METAL предоставляет идеальные условия для организации эффективной вентиляции пространства под кровлей. Пленка способна обеспечить непрерывную циркуляцию потоков воздуха, в результате чего влажный воздух, конденсат и пар максимально быстро отводятся из чердачного помещения. Более того, использование этой супердиффузионной мембранной пленки позволяет создать между крышей и утеплителем определенное термическое пространство, благодаря которому в летнее время года уменьшается нагрев дома, в зимний период на поверхности крыши не будет появляться наледь.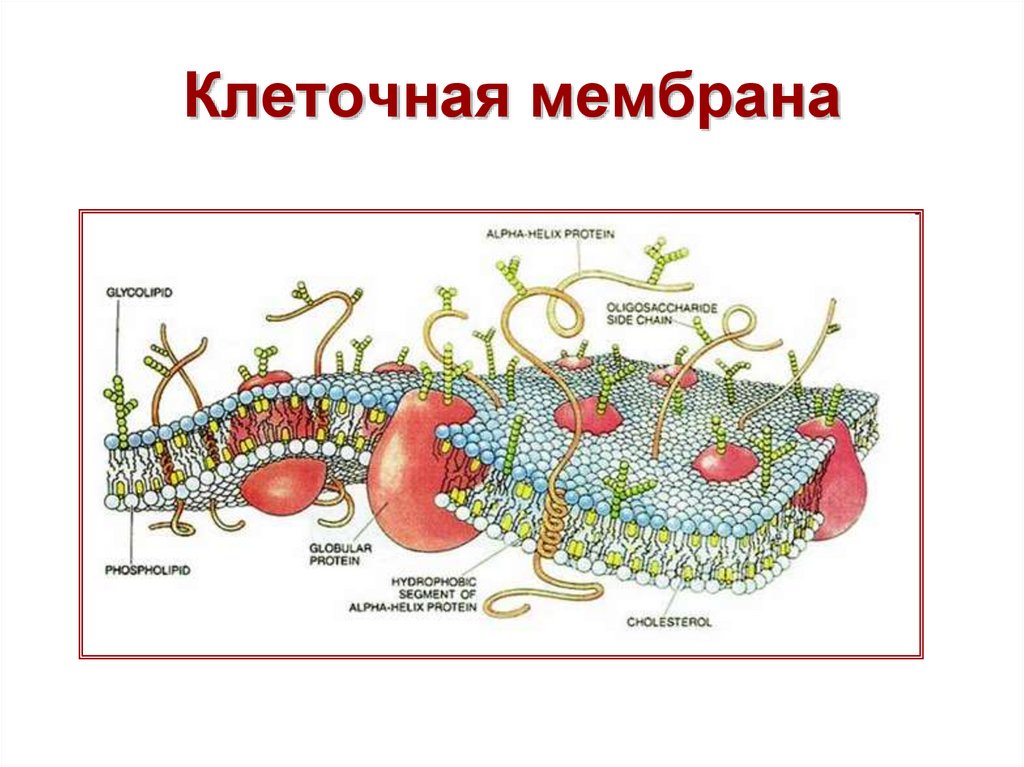 Таким образом, в настоящее время на рынке строительных материалов присутствует множество самых разнообразных супердиффузионных мембран, обладающих различными характеристиками. Выбор того или иного типа зависит от конкретных условий эксплуатации и климатической зоны, в пределах которой расположено строение.
Таким образом, в настоящее время на рынке строительных материалов присутствует множество самых разнообразных супердиффузионных мембран, обладающих различными характеристиками. Выбор того или иного типа зависит от конкретных условий эксплуатации и климатической зоны, в пределах которой расположено строение.
Инстаграм
- 0,0498 s
- ©2018 Все права защищены
superarch.ru
Изоляционные стройматериалы: плёнки и диффузионные мембраны
Несмотря на развитие строительных технологий, скатные крыши остаются основным способом покрытия здания, одновременно простым и эффективным. Но во многих домах чердак перестал быть просто чердаком, а стал функциональным помещением, пригодным для жилья. Для того чтобы превратить чердаки из заброшенных кладовых, где собирались старые вещи, в жилые помещения, мастерские, офисы, их необходимо гидроизолировать, утеплить, организовать освещение и вентиляцию. В связи с этим старые технологии устройства подкровельного пространства претерпели определённые изменения, а на смену старым материалам пришли новые, обладающие уникальными свойствами.
Необходимость применения изоляционных плёнок и мембран
Для решения задач по устройству мансардных помещений всё более востребованными становились подкровельные плёнки. Причины их появления:
- Необходимость гидроизоляции кровли, вызванная постоянным воздействием на неё атмосферных явлений.
- Кровельные покрытия являются не очень эффективными теплоизоляторами, поэтому мансарды подвергаются сильному температурному воздействию. Кроме того, снег, ветер и дождь ещё более снижают теплоизолирующую эффективность кровли. Поэтому в конструкции кровельного покрытия мансарды должен присутствовать надёжный теплоизолятор, защищённый от проникновения влаги.
- Необходимость создания звукоизолирующего слоя, не пропускающего внутрь подкровельного пространства шум дождя, ветра и прочие посторонние звуки.
- Использование человеком большого количества бытовой техники, нагревающей воздух и испаряющей влагу. По законам физики пар поднимается вверх, конденсируясь на потолке или проникая в элементы конструкции крыши и разрушая их.

Для сохранения необходимого влажностного режима внутри здания, предотвращения намокания конструктивных элементов крыши, в том числе утеплителя, используют подкровельные плёнки и мембраны.
Подкровельные плёнки: виды и их характеристики
Подкровельные плёнки представляют собой двух- или трёхслойный материал, прозрачный или серого цвета, представляющий собой полиэтиленовое полотно, как вариант – армированное полипропиленовой сеткой. Полиэтиленовое полотно обеспечивает хорошие гидроизолирующие характеристики материала, а полипропиленовый армирующий слой – прочность на растяжение и разрыв. Плёнки бывают перфорированными и не перфорированными, что определяет назначение материала.
Основным назначением паро- и гидроизоляционных плёнок является обеспечение сохранения рабочих характеристик кровельной теплоизоляции. Эти материалы предохраняют теплоизоляцию от проникновения в неё влаги, что может привести не только к снижению её эффективности, но и полному разрушению.
Виды подкровельных плёнок:
- Пароизоляционные плёнки применяют при устройстве тёплых чердачных помещений. Такие плёнки предназначены для предохранения нижнего слоя утеплителя от попадания в него водяных паров, образующихся в нижних помещениях. Этот материал является барьером для поднимающейся влаги, удерживая её до момента вентиляции помещения.
- Гидроизоляционные плёнки (на фото справа) сочетаются с любым типом кровельных материалов и предназначаются для предохранения от влаги верхнего слоя утеплителя. Этот тип плёнок, благодаря микроперфорации, имеет более высокую паропроницаемость, чем пароизоляционные плёнки,. При монтаже гидроизоляционной плёнки между нею и теплоизоляцией необходимо предусматривать зазор, предназначенный для удаления влаги из утеплителя и предотвращения образования конденсата на плёнке. Для холодных чердачных помещений применение таких плёнок эффективно и оправдано при простой конструкции крыши. Для сложных конфигураций подкровельного пространства устройство зазора является затруднительным.
 При выборе гидроизоляционных плёнок основным параметром является водонепроницаемость, измеряемая в мм водяного столба. Чем выше этот показатель, тем эффективней материал. Учитываются также такие характеристики, как прочность и стойкость к ультрафиолетовому излучению.
При выборе гидроизоляционных плёнок основным параметром является водонепроницаемость, измеряемая в мм водяного столба. Чем выше этот показатель, тем эффективней материал. Учитываются также такие характеристики, как прочность и стойкость к ультрафиолетовому излучению. - Антиконденсатные плёнки применяют при устройстве металлических кровель, к которым относятся фланцевые кровельные покрытия, металлочерепица, профнастил, еврошифер. Для таких кровель большую опасность представляет влага, которая конденсируется на нижней поверхности листа. Она вызывает коррозию и приводит к разрушению кровельного материала. Одна сторона плёнки изготовлена из ворсистого нетканого текстиля, прекрасно впитывающего влагу, а другая – из полиэтиленовой плёнки. Между плёнкой и утеплителем предусматривается зазор, благодаря которому выветривается задержанная влага.
Особенности мембран
Мембраны являются относительно новым материалом. В отличие плёнок, которые использовались ранее, строительные мембраны благодаря своей нетканой структуре выпускают наружу весь пар, находящийся в помещении, но не дают просочиться внутрь конструкции крыши влаге, которая просачивается через кровлю. Примером природного материала, обладающего способностью выпускать влагу наружу, не пропуская её внутрь, является кожа.
Примером природного материала, обладающего способностью выпускать влагу наружу, не пропуская её внутрь, является кожа.
При монтаже подкровельных мембран организация дополнительного зазора не требуется, так как этот материал не создаёт сопротивление для водяного пара. Укладывать изоляционную мембрану можно непосредственно на утеплитель.
Мембраны отличаются между собой технологиями производства, применяемым нетканым материалом, наличием специальных добавок, увеличивающих УФ-стабильность, количеством слоёв, вида водонепроницаемых плёнок и типа их скрепления.
Этот строительный материал широко применяется при строительстве зданий с отапливаемыми чердаками и мансардами.
Мембраны различаются по способу укладки. Они бывают:
- одностороннего применения, укладываются поперёк стропил определённой стороной наружу;
- двустороннего применения, этот материал можно укладывать на утеплитель любой стороной.
Основные показатели, которые необходимо учитывать при выборе мембран
Существует несколько критериев, служащих для выбора мембраны для конкретных эксплуатационных условий.
Основная техническая характеристика – паропроницаемость. Коэффициентом паропроницаемости называют количество пара, пропускаемое за сутки мембраной площадью 1м2 при разности парциального давления по обе стороны мембраны равной 1 МПа, и одинаковой температуре. Измерение этого параметра осуществляют при комнатной температуре и максимальной разнице влажности по обе стороны мембраны. Таким способом определяют идеальную, то есть, максимальную паропроницаемость материала. Понятие «коэффициент паропроницаемости» часто заменяют понятием «паропроницаемость». Для эффективного удаления влаги из утеплителя эта величина должен составлять не менее 400 г/м2 в сутки. Коэффициент паропроницаемости ниже 300 г/м2 в сутки считается недостаточным. По этому критерию мембраны разделяют на следующие виды:
- псевдодиффузионные – менее 300 г/м2 в сутки;
- диффузионные – 400-1000 г/м2 в сутки;
- супердиффузионные – более 1000 г/м2 в сутки.
На практике вполне приемлемым является материал с коэффициентом паропроницаемости 600 г/м2 в сутки.
Вторым показателем паропроницаемости мембран служит сопротивление паропроницаемости. Эта величина равна разности парциального давления по обе стороны мембраны, при которой сквозь 1м2 материала за час проходит 1 мг водяного пара, при равной температуре с противоположных сторон.
Для сравнения сопротивления проникновению пара мембраны и воздуха служит коэффициент сопротивления диффузии пара. Он показывает, во сколько раз сопротивление мембраны диффузии через неё водяного пара выше, чем сопротивление слоя сухого воздуха такой же толщины.
Основные характеристики диффузионных мембран
Диффузионные мембраны служат для защиты подкровельного пространства от влаги. Представляют собой полиэтиленовые плёнки, армированные перлинковой нитью. Диффузионная мембрана, в отличие от гидроизоляционной плёнки, пропускает пар в верхний вентиляционный зазор, надёжно защищая, при этом, подкровельное пространство от проникновения пыли и влаги снаружи.
Эти мембраны не рекомендованы к применению в сочетании с металлическим кровельным покрытием, поскольку они свободно пропускают пар из помещения, что приводит к разрушению металла из-за коррозии. Применение диффузионных мембран требует устройство только верхнего вентиляционного зазора, что позволяет значительно сэкономить материалы, из которых пришлось бы сооружать дополнительную обрешётку. Такие мембраны часто используют для кровель сложной конфигурации.
Применение диффузионных мембран требует устройство только верхнего вентиляционного зазора, что позволяет значительно сэкономить материалы, из которых пришлось бы сооружать дополнительную обрешётку. Такие мембраны часто используют для кровель сложной конфигурации.
К материалам данной группы относятся мембраны Tyvek Soft c коэффициентом паропроницаемости 744 г/м2 в сутки и Изоспан АМ с паропроницаемостью 850 г/м2. Трёхслойная диффузионная мембрана Delta Vent изготовлена из комбинации паропроницаемых мембран из нетканого полипропиленового полотна. Материал обладает повышенной прочностью, стойкостью к механическим повреждениям во время проведения кровельных работ.
Особенности супердиффузионных мембран
Супердиффузионные мембраны предназначены для надёжной защиты подкровельного пространства от дождя и снега. Высокая паропроницаемость этого материала обеспечивает эффективное выведение влаги из утеплителя. Как и для диффузионных, для супердиффузионных мембран требуется только один верхний вентиляционный зазор. Оба этих материала могут применяться с битумной, керамической, цементнопесчаной черепицей, композитной металлочерепицей.
Оба этих материала могут применяться с битумной, керамической, цементнопесчаной черепицей, композитной металлочерепицей.
Выбор супердиффузионной мембраны является прекрасным вариантом при переоборудовании чердачного помещения в мансарду без перестройки стропильной конструкции.
К супер диффузионным относятся трёхслойные мембраны Евробарьер (на фото слева), включающие решётку из полипропиленовых волокон, ламинированную с двух сторон полиэтиленом. В материале присутствуют специальные стабилизирующие добавки, которые придают ему повышенную стойкость к воздействию солнечных лучей. Это позволяет немного задержать проведение работ по укладке кровельного покрытия.
Объёмные разделительные диффузионные мембраны
Этот вид мембран монтируют с одним верхним вентиляционным зазором на кровлях с металлическим покрытием – из меди, стали, алюминия, цинк-титанового сплава. Рекомендован для применения на кровлях сложной конфигурации – с куполами, башнями, слуховыми окнами.
Объёмная диффузионная мембрана Delta Trela эффективна для применения на крышах с пологими скатами – с уклоном до 150, где велика вероятность скопления конденсата с внутренней стороны листа.
Технологии производства подкровельных плёнок и мембран постоянно развиваются и предлагают потребителю всё более совершенные гидро- и пароизоляционные материалы. Использование подкровельных плёнок и мембран позволяет повысить надёжность и долговечность конструкции крыши, тем самым избавив потребителя от частых дорогостоящих ремонтов.
GD Star Ratingloading…
Диффузионные мембраны: основные характеристики и особенности их применения, 3.2 out of 5 based on 14 ratings
izolyar.com
Что это и какие бывают, инструкция по монтажу, цены за рулон
Кровельная мембрана — современный материал, осуществляющий гидроизоляцию кровельного пирога от осадков, конденсата и скапливания пара в утеплителе. В статье вы узнаете для чего используется кровельная мембрана, какие виды существуют и как сделать правильный монтаж мембраны.
Для чего нужна кровельная мембрана
Кровельная мембрана используется в следующих случаях:
- Если есть разность температур между этажами — например, если мансарда зимой не отапливается или отапливается изредка;
- Если есть разность температур между внутренней и внешней обшивкой — например, при отапливаемом чердаке в холодное время года;
- Если используются металлические кровельные материалы — из-за перепадов температур на внутренней поверхности кровли образуется конденсат, разрушающий покрытие.

Кровельная мембрана выпускает лишний пар наружу, при этом защищает от попадания воды в дом (например, от осадков). Единственное условие использования мембран — хорошая вентиляция. Без свободной циркуляции воздуха мембраны неэффективны, поэтому еще на этапе монтажа крыши нужно предусмотреть все технологические зазоры между утеплителем, гидроизоляцией и кровлей, требуемые производителем.
Ни в коем случае нельзя укладывать обычную гидроизоляцию (не пропускающую пар) на перекрытия, которые разделяют теплые и холодные помещения — в утеплителе неизбежно будет образовываться конденсат. Как результат, мокрый утеплитель не только перестает выполнять свои функции, но и становится источником плесени и грибков.
Виды кровельных мембран
Сфера применения мембран чрезвычайно широка — от изоляции внутренних перегородок до устройства крыши бани. По своему типу они делятся на:
Изготавливаются мембраны из нетканого полотна из полимерных волокон. Своими уникальными качествами мембраны обязаны многослойной структуре — от двух до четырех слоев.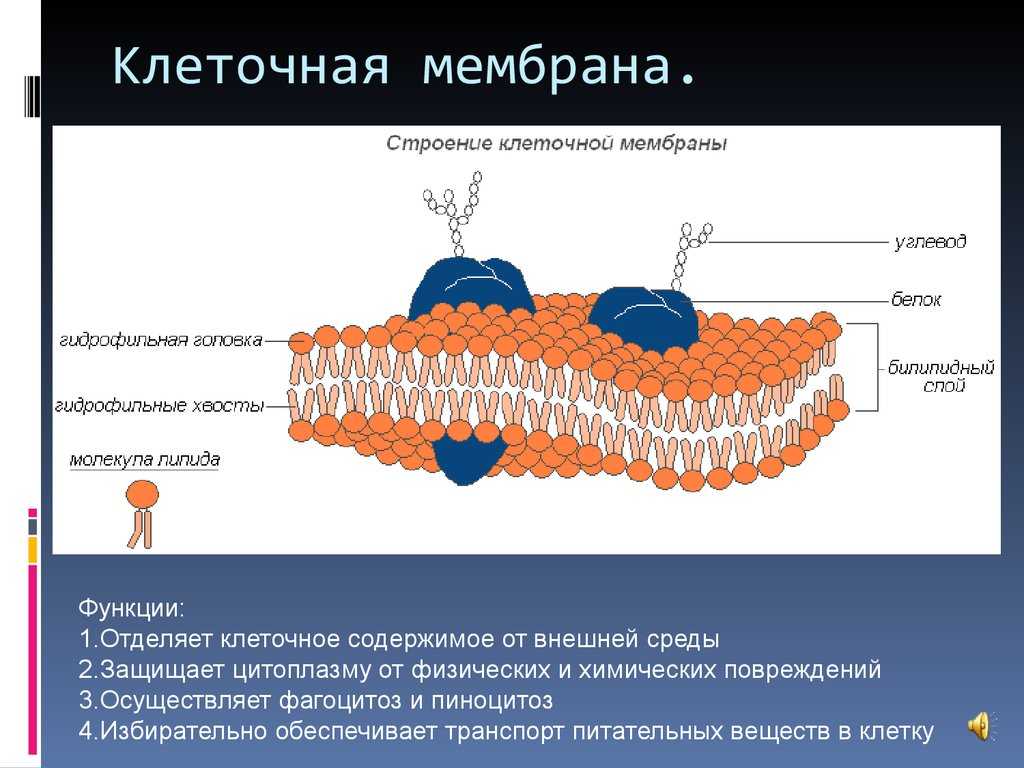 Верхний обеспечивает УФ-защиту, нижний позволяет беспрепятственно проникать пару, а внутренний выполняет функцию барьера, задерживая влагу и не пропуская её в нижний слой.
Верхний обеспечивает УФ-защиту, нижний позволяет беспрепятственно проникать пару, а внутренний выполняет функцию барьера, задерживая влагу и не пропуская её в нижний слой.
Диффузионная мембрана
Диффузионная мембрана обладает паропроницаемостью до 1000 мг/м², которой достаточно для использования с практически любыми кровлями в умеренном климате. Показатель паропроницаемости указывает на способность материала пропускать заданное количество влаги в сутки на один квадратный метр. Подробнее.
Супердиффузионная мембрана
Супердиффузионная мембрана отличается еще большей паропроницаемостью — от 1000 мг/м². Применяется она в широтах с повышенной влажностью, большими перепадами температур или для утеплителей большой толщины. Они отлично зарекомендовали себя в каркасном строительстве, где требования к состоянию утеплителя существенно выше. Подробнее.
Антиконденсатная мембрана
Если в качестве кровельного материала для дачи используется металлочерепица или профнастил, антиконденсатная мембрана является важнейшим элементом кровельного пирога. Благодаря уникальной двухслойной структуре, пар не испаряется под металлическое покрытие и не способствует его разрушению. Нижний ворсистый слой впитывает влагу, которая просто стекает наружу, а верхний защищает утеплитель от попадания конденсата и осадков на утеплитель.
Подробнее.
Благодаря уникальной двухслойной структуре, пар не испаряется под металлическое покрытие и не способствует его разрушению. Нижний ворсистый слой впитывает влагу, которая просто стекает наружу, а верхний защищает утеплитель от попадания конденсата и осадков на утеплитель.
Подробнее.
Кровельные мембраны Ондутис
Благодаря своим свойствам мембраны Ондутис являются одними из самых популярных гидроизоляционных материалов.
Мембраны обладают следующими характеристиками:
| Устойчивость к атмосферным воздействиям | Уложенная на стропила мембрана может находиться под открытым небом больше месяца без потери своих качеств |
| Температурный диапазон от -40 до +80 | Мембраны могут укладываться в любое время года, а сильное переохлаждение или нагрев не влияют на гидроизоляционные свойства |
| Водоупорность больше 1000 мм водного столба | Даже затяжной проливной дождь не сможет проникнуть под такую мембрану |
| Устойчивость к разрыву | Супердиффузионные мембраны Ондутис могут использоваться как временная кровля на срок до 2х месяцев |
Как выбрать мембрану
Выбирая мембрану, важно заранее определиться с типом кровли, временем проживания (теплый дом или холодный) и схемой отопления. Так, для деревянного дома, где будет отапливаться только первый этаж, для крыши можно использовать обыкновенную гидроизоляцию. А вот со стороны чердачного перекрытия лучше уложить мембрану с паропроницаемостью до 300 мг/м² — это защитит утеплитель от образования конденсата.
Так, для деревянного дома, где будет отапливаться только первый этаж, для крыши можно использовать обыкновенную гидроизоляцию. А вот со стороны чердачного перекрытия лучше уложить мембрану с паропроницаемостью до 300 мг/м² — это защитит утеплитель от образования конденсата.
Для летнего дачного домика паропроницаемую мембрану можно заменить рулонной гидроизоляцией, существенно снизив стоимость строительства. А вот при устройстве бани или сауны из бруса кроме супердиффузионной мембраны с внешней стороны рекомендуется использовать фольгированную отражающую пароизоляцию (например: R Termo). Это не только продлевает срок службы кровли, но и обеспечивает отсутствие конденсата в помещении.
Монтаж кровельной мембраны: пошаговая инструкция
Процесс монтажа довольно прост, единственное условие — соблюдать технические зазоры и нахлесты, рекомендуемые производителем:
- Укладка мембраны производится поверх стропил вплотную к утеплителю. Рулоны раскатываются горизонтально и начинают крепиться снизу вверх — от карниза к коньку.
- Если конструкция крыши включает ендовы или косые коньки, предварительно вдоль них прокладываются дополнительные полосы, а основная гидроизоляция укладывается поверх.
- К стропилам изоляция может крепиться как строительным степлером, так и оцинкованными гвоздями с широкой шляпкой.
- Между собой пленка соединяется специальной монтажной лентой с вертикальным нахлестом в 20 см и горизонтальным в 15 см. Вертикальные стыки должны обязательно находиться над стропилами — их дополнительно закрепляют степлером.
- Фиксируется мембрана брусками контробрешетки высотой минимум 5 см, закрепляемыми вдоль стропил — это обеспечит достаточное пространство для вентиляции.
- Для укладки мембраны вокруг дымохода или вентиляции, в мембране прорезается отверстие в виде буквы Н, куда и продевается проходной элемент. Края фиксируются на дымоходе монтажной лентой так, чтобы обеспечить полноценную гидроизоляцию.

- Мембраны Ондутис не рекомендуется заводить в водосточный желоб — нижняя кромка просто закрепляется на капельнике.
Также смотрите наше видео об укладке кровельной мембраны.
1 голос , пожалуйста, оцените статью:
ondutis.ru
Что такое диффузионная мембрана
Вступление
Важнейшим слоем в «пироге» многослойной конструкции кровли является слой гидроизоляции. Лучшим материалом для гидроизоляции кровли является диффузионная мембрана. Данный строительный материал относится к группе подкровельные пленки и мембраны.
Диффузионная мембрана это
Диффузионная мембрана – материал, используемый для гидроизоляции кровли, предохранения от попадания влаги в утеплитель и производства других работ в строительстве. Данный продукт новых технологий появился сравнительно недавно. От обыкновенных пленок (использовавшихся ранее с той же целью) отличается своей способностью к однонаправленному пропусканию влаги. При этом не пропускает ни воду, ни воздух, работая одновременно и тепло и гидроизолятором.
Устройство диффузионной мембраны
Данный материал предлагается в виде рулонов (для удобства использования). Многослойное полотно, представляющее собой капиллярный насос (влага перемещается с ворсистой поверхности к гладкой). Мембраны Ондутис (продукция компании Ондулин) 3-слойные – снаружи нетканый полипропилен, внутри стойкий к растяжению усилитель. Благодаря такой конструкции, мембрана этой марки имеет не только высокие характеристики гидрофобности и паропроницаемости, но и отличную механическую прочность. Слои мембраны соединяются в пакет посредством высокотехнологичной ультразвуковой сварки. Материал экологически безопасен и устойчив к воздействию микроорганизмов.
Слои мембраны соединяются в пакет посредством высокотехнологичной ультразвуковой сварки. Материал экологически безопасен и устойчив к воздействию микроорганизмов.
Свойства диффузионной мембраны
Важнейшим свойством диффузионной мембраны является её паропроницаемость. Измеряется она в миллиграммах пара, проходящих через квадратный метр поверхности за 24 часа. В зависимости от ее величины различают мембраны:
- Мало диффузионные. Менее 300 мг/м2/сутки. Используются только внутри помещений.
- Средне диффузионные. 300-1000 мг/м2/сутки. Наиболее массовые в использовании. Отлично работают в зонах с умеренным климатом.
- Высоко диффузионные (супердиффузионные). Свыше 1000 мг/м2/сутки. Используются при строительстве в сложных климатических условиях (или при использовании толстого утеплителя), в местах с резкой сменой влажности.
С какой целью разработана диффузионная мембрана
Материал был разработан для защиты утеплителя от промокания (и как следствие, потери теплоизоляционных свойств), сохранения в рабочем состоянии стропил, и других внутренних элементов кровли, защиты стен и сохранения тепла в доме, в целом.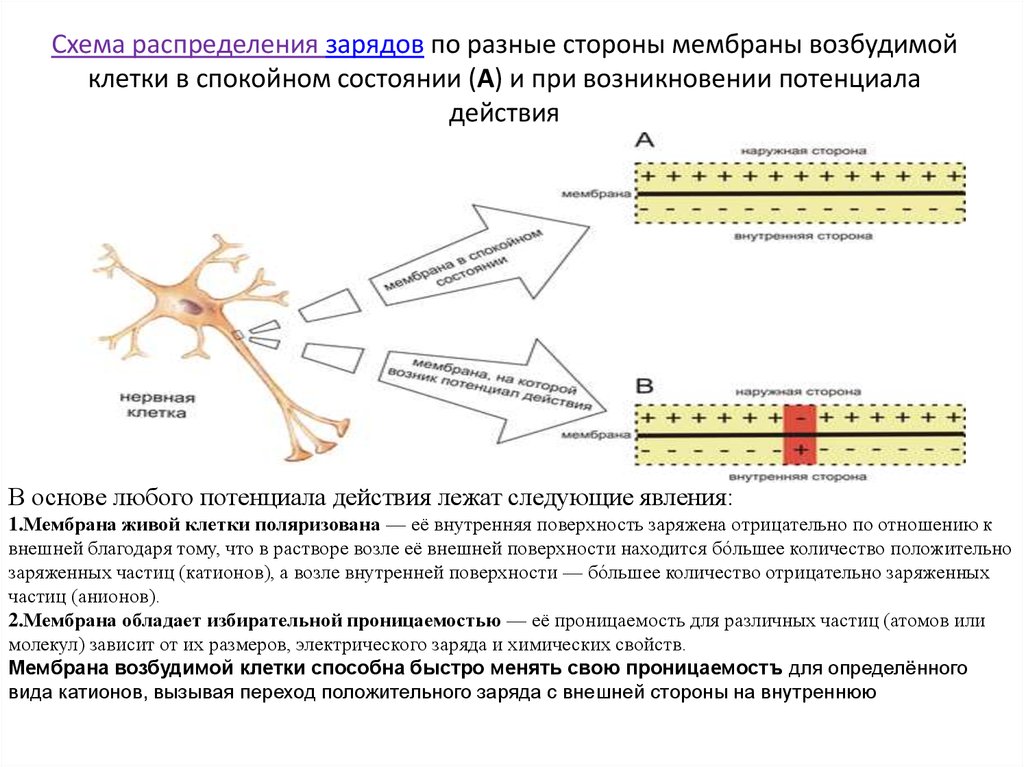 Материал воздухопроницаем, благодаря чему дом дышит, и не нуждается в дополнительном проветривании. В то же время не пропускает воду, защищая жилище от непогоды. Благодаря высокой паропроницаемости происходит быстрое удаление водного конденсата из-под кровли или наружной облицовки стен.
Материал воздухопроницаем, благодаря чему дом дышит, и не нуждается в дополнительном проветривании. В то же время не пропускает воду, защищая жилище от непогоды. Благодаря высокой паропроницаемости происходит быстрое удаление водного конденсата из-под кровли или наружной облицовки стен.
Мембрана обладает свойством односторонней проницаемости. Вода забирается от утеплителя, затем по другой стороне стекает или испаряется. С учетом этого при производстве стороны красят в разный цвет. А на одну из сторон наносятся надписи, пиктограммы, фирменные изображения. Ошибка в выборе правильной ориентации материала при монтаже приведет к тому, что теплоизоляция намокнет, и ее свойства резко ухудшатся.
Где используется диффузионная мембрана
Мембраны массово применяются в строительстве зданий и сооружений для гидроизоляции крыш (при наличии подкровельной вентиляции), теплоизоляции и влагозащиты стропил. Защищают несущие деревянные конструкции. Используют в вентиляционных навесных фасадах. При использовании такого материала в деревянном домостроении НЕ требуется дополнительная пропитка древесины антисептиком.
При использовании такого материала в деревянном домостроении НЕ требуется дополнительная пропитка древесины антисептиком.
Кроме того, диффузионная мембрана применяется при обустройстве не утепленной кровли из:
- полимерной или керамочерепицы;
- металлочерепицы;
- шифера;
- ондулина.
Сильные стороны диффузионных мембран
К основным преимуществам данной продукции можно отнести:
- высокую механическая прочность;
- температурную выносливость (хорошо выносит перепады температуры), при нагреве – не выделяет опасных элементов;
- устойчивость к химически активным реагентам;
- экологичность.
Тонкости укладки диффузионной мембраны
Укладка полотна обязательно проводится с учетом направления работы мембраны. Продукция Ондутис кладется логотипом наружу, поверхностью без надписей – к утеплителю. Все стыки и неплотные прилегания должны быть надежно заизолированы, во избежание проникновения воды в утеплитель. Необходимо тщательно изолировать места выходов труб, антенн и других проникающих конструкций. В мембране под них производится трапецеидальный надрез.
Необходимо тщательно изолировать места выходов труб, антенн и других проникающих конструкций. В мембране под них производится трапецеидальный надрез.
Для нормальной работы мембраны, надо заблаговременно обеспечить естественную вентиляцию под кровлей с целью отвода водяного пара, создать продуваемый промежуток между ней и покрытием. Размер зазора обычно составляет 70-120мм (сумма толщин брусков обрешетки и контррейки). Для доступа наружного воздуха предусматривают вентиляционные отверстия.
Если мембрана используются для утепления стен снаружи, ее монтируют прямо к утеплителю. Крепление мембраны производится в строительный каркас (сквозь теплоизоляцию) скобами или гвоздями. Поверх устанавливаются направляющие, к которым крепится облицовка. В итоге между облицовкой и пленкой создается необходимый вентиляционный зазор.
Супердиффузионная мембрана – что это и зачем нужна
Инновационный продукт, созданный для условий с экстремальными температурами и влажностью, назначение, которого – эффективная защита теплоизоляции с высокими показателями паропроницаемости.
Новые продукты – супермембраны Ондутис SA115 и SA130 еще лучше пропускают пар, но обеспечивают надежную преграду воде и воздуху. Надежно оберегают теплоизоляцию и конструктивные элементы зданий, которые не переносят влажность. Помогая сохранять свойства теплозащиты, они снижают затраты на поддержание тепла в доме.
©DomiTy.ru
Статьи по теме
Какую выбрать супердиффузионную мембрану?
- ПОСЛЕДНИЕ ЗАПИСИ Краски и эмали по металлу и ржавчине Пленка гидроизоляционная для крыши Как выбрать хорошую акриловую ванну Какие бывают розетки, их устройство, типы и классификация
- РУБРИКИ
- Автоматическое открывание, проветривание и полив теплиц
- Акриловые краски
- Балкон
- Блоки арболитовые
- Бурение скважин на воду
- Вода из скважины
- Водосток кровельный
- Воздух в квартире
- Выращивание дома
- Гидроизоляция
- Гидрофобизация материалов
- Дорожки садовые
- Камин своими руками
- Каркасный дом
- Кладка печи своими руками
- Крыша из металлочерепицы
- МДФ
- Монтаж кровли
- Монтаж ламината
- Монтаж линолеума
- Монтаж подложки под ламинат
- Натяжные потолки
- Опилкобетон
- ОСБ плита
- Отделка откосов
- Оштукатуривание
- Полипропиленовые трубы
- Расход материалов
- Тротуарная плитка
- Устройство отмостки
- Утепление
- Утепляем баню самостоятельно
- Фасад
- Фундамент из свай
- Шлакоблок
- Эмаль для ванны
Как известно, любая кровельная конструкция представляет собой многослойную структуру, в которой слой утеплителя должен быть обязательно защищен от проникновения влаги паро- и гидроизоляционными пленками. Какими характеристиками должны обладать пленки, предназначенные для защиты утепляющего слоя, учитывая, что даже при абсолютной целостности крыши влага все равно попадает в подкровельное пространство?
Какими характеристиками должны обладать пленки, предназначенные для защиты утепляющего слоя, учитывая, что даже при абсолютной целостности крыши влага все равно попадает в подкровельное пространство?
Что такое супердиффузионные мембраны?
Диффузионная мембрана – это специальный материал, имеющий двух-, трех- или даже четырехслойную структуру, основу которого составляет нетканый холст. Диффузионные мембраны применяют для защиты утепляющего слоя от проникновения в его толщу испарений. Также, диффузионные мембраны являются превосходной защитой от воды и ветра.
Также, диффузионные мембраны являются превосходной защитой от воды и ветра.
При создании крыши, в полном объеме соответствующей всем современным требованиям, каждый застройщик обязательно столкнется с таким понятием, как «кровельный пирог». Для того чтобы крыша выполняла все возложенные на нее функции в течение всего срока эксплуатации, кроме основного кровельного покрытия, необходимо использовать некоторые дополнительные материалы, к числу которых относятся супердиффузионные мембраны.
Супердиффузионные мембраны можно использовать при создании кровельного пирога в любой климатической зоне нашей страны. Роль этого дополнительного слоя чрезвычайно важна, так именно его присутствие позволяет снизить силу неблагоприятных воздействий, вызванных экстремальными погодными условиями, а также нивелировать недочеты и ошибки, возникшие в ходе неправильного монтажа кровли.
Предназначение диффузионных мембран
Основное предназначение супердиффузионных мембран для кровли является обеспечение защиты от проникновения внутренней и наружной влаги внутрь теплоизоляционного слоя.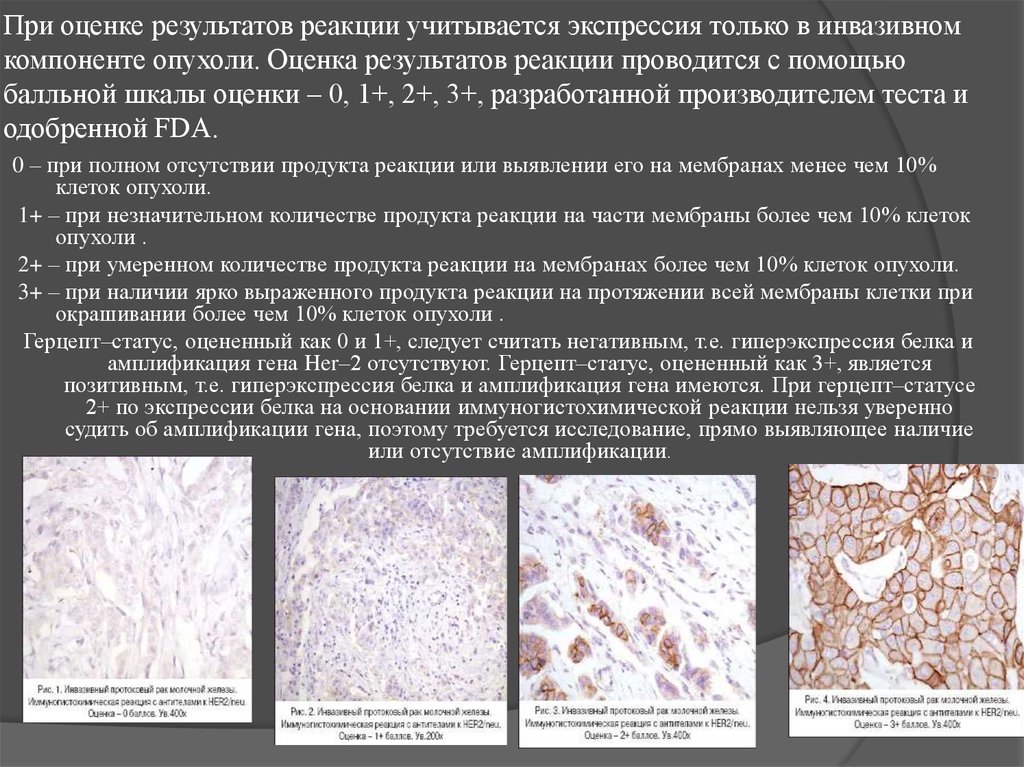 Источниками этой влаги могут быть внутренние испарения и атмосферные осадки. Кроме этого, расположенная в кровельном покрытии диффузионная мембрана обеспечивает эффективные условия отвода уже накопившейся в силу тех или иных причин влаги.
Источниками этой влаги могут быть внутренние испарения и атмосферные осадки. Кроме этого, расположенная в кровельном покрытии диффузионная мембрана обеспечивает эффективные условия отвода уже накопившейся в силу тех или иных причин влаги.
Супердиффузионную мембрану можно с полной уверенностью назвать одной из важнейших составляющих теплоизоляционного контура, так как она косвенным образом способствует снижению потерь тепловой энергии. Бережливый хозяин собственного дома, знающий толк в экономии, никогда не будет раздумывать о необходимости или отсутствии таковой при принятии решения о покупке и последующей установке диффузионной мембраны. Тем более, что стоимость этого материала на современном рынке строительных материалом можно с уверенностью назвать чисто символической.
Преимущества использования супердиффузионных мембран
Хозяин частного дома, решивший использовать в конструкции кровельного пирога супердиффузионные мембраны, в сравнении с домовладельцами, использующими традиционные технологии, получит ряд неоспоримых преимуществ, среди которых основными можно назвать следующие:
- Использование супердиффузионных мембран позволяет одной пленке заменить две, такие как гидро- и ветрозащита.
 Наличие мембраны допускает возведение конструкции без наличия вентиляционного зазора.
Наличие мембраны допускает возведение конструкции без наличия вентиляционного зазора. - Укладка супердиффузионных мембран разрешается непосредственно на поверхность любого покрытия, что позволяет укладывать теплоизоляцию более толстым слоем, в сравнении с традиционными технологиями. Как результат, владелец дома получает усиленную теплоизоляцию.
- Использование супердиффузионных мембран позволяет продлить срок эксплуатации утепляющего материала и деревянных конструкций кровли. При этом, деревянные элементы крыши могут быть установлены без предварительной обработки специальными химическими составами.
- Применение супердиффузионных мембран в ходе создания кровельного пирога значительно сокращает время проведения монтажных работ и связанных с ними затрат.
Разновидности супердиффузионных мембран
В свободной продаже супердиффузионные мембраны можно встретить в виде рулонов. Рулоны имеют незначительную массу, благодаря чему проведение работ, связанных с благоустройством кровли, проходит достаточно просто. Диффузионные мембраны обладают следующими основными характеристиками: количество слоев, степень проницаемости пара, плотность материала, водопроницаемость. Необходимый для конкретных условий материал необходимо выбирать на основе числовых значений этих показателей.
Рулоны имеют незначительную массу, благодаря чему проведение работ, связанных с благоустройством кровли, проходит достаточно просто. Диффузионные мембраны обладают следующими основными характеристиками: количество слоев, степень проницаемости пара, плотность материала, водопроницаемость. Необходимый для конкретных условий материал необходимо выбирать на основе числовых значений этих показателей.
В настоящее время на рынке присутствуют следующие разновидности супердиффузионных мембран:
1. Difoil-S. Диффузионная пленка этого типа имеет трехслойную структуру. Материал характеризуется высокой степенью паропроницаемости, он способен достаточно быстро отводить из помещения водяной пар. Чаще всего эту пленку укладывают непосредственно на теплоизолирующий материал или на поверхность настила, при этом необходимость создания вентиляционного зазора отсутствует. Максимально успешно данная супердиффузионная пленка может быть использована для мансардных крыш, она превосходно сочетается с обыми типами кровельных материалов, присутствующих в настоящее время на строительном рынке.
Difoil-S способен обеспечить эффективную защиту крыши и стен от влаги и ветровых нагрузок. Чаще всего, этот материал используют в ходе утепления наружных стен. В вентилируемых кровельных конструкциях данную диффузионную пленку укладывают непосредственно на поверхность теплоизоляционного материала, что позволяет создать дополнительную защиту от проникновения атмосферной влаги. Difoil-S превосходно сочетается с любыми типами изолирующих материалов. Монтируется данная мембрана с небольшим провисанием или сверху слоя теплоизоляции стороной, на которой имеются надписи, в направлении крыши.
2. MASTERMAX 3 ECO – супердиффузионная мембрана для подкровельного применения, характеризующаяся высоким качеством и превосходными эксплуатационными характеристиками. Этот материал используют в качестве защитного покрытия кровельной конструкции. Его использование позволяет создать надежную защиту утеплителя от влаги, ветра и пыли. Также, материал способен отводить от поверхности теплоизолятора испарения.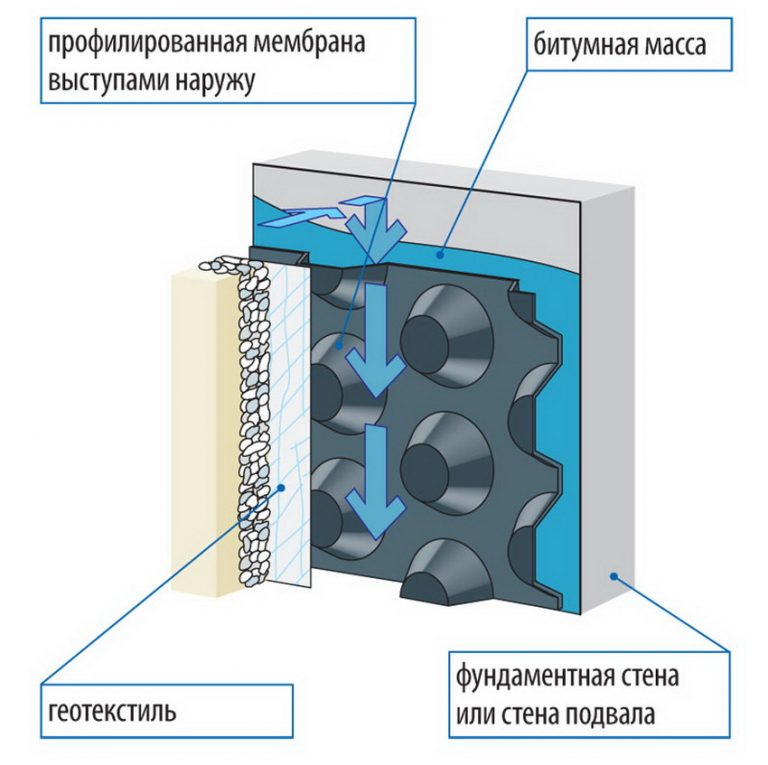
В виду того, что диффузионная мембрана изготовлена на основе полипропиленового волокна, она обладает высокой паропроницаемостью и увеличенной способностью пропускания водяных паров из пространства под кровлей. Благодаря ниличию подробных характеристик, MASTERMAX называют мембранами новейшего поколения.
Мембраны MASTERMAX 3 ECO могут эксплуатироваться с любыми видами теплоизоляционных материалов. Для получения максимальной эффективности, мембрану необходимо устанавливать правильно, а именно, нанесенная на поверхность материала надпись MASTERMAX 3 ECO должна быть расположена в сторону кровли.
3. MASTERMAX 3 CLASSIC. Данный материал представляет собой трехслойную гидроизоляционную супердиффузионную мембрану. Он используется для создания второстепенной защиты от воздействия влаги и пыли. Пленку монтируют сверху теплоизоляционного слоя.
4. MASTERMAX 3 TOP. Данная супердиффузионная мембрана предназначена для защиты слоя утеплителя тех кровель, которые эксплуатируются в условиях повышенных ветровых и влажностных нагрузок. Эту пленку, также как и прочие ее аналоги, устанавливают поверх слоя утеплителя.
Эту пленку, также как и прочие ее аналоги, устанавливают поверх слоя утеплителя.
5. MASTERMAX 3 EXTRA представляет собой высокотехнологичную мембрану, которую также как и MASTERMAX 3 TOP монтируют в пределах кровельного пирога крыш, подверженных экстремальным влажностным и ветровым нагрузкам. Данная разновидность мембран является наиболее плотной и самой высококачественной пленкой среди представителей трехслойных мембран от производителя MASTERMAX. Кроме своего основного предназначения, пленка MASTERMAX 3 EXTRA способна успешно противостоять воздействию прямых ультрафиолетовых лучей. Данную мембрану укладывают без вентиляционного зазора.
6. MASTERMAX METAL – это пленка, которая относится к категории четырехслойных супердиффузионных мембран. Этот паропроницаемый материал используют в конструкции кровельного пирога, в котором он выполняет роль второстепенной защиты от снега, дождя, тумана, ветра и пыли. MASTERMAX METAL укладывают под металлические листы жестяных крыш, лежащих на деревянной основе.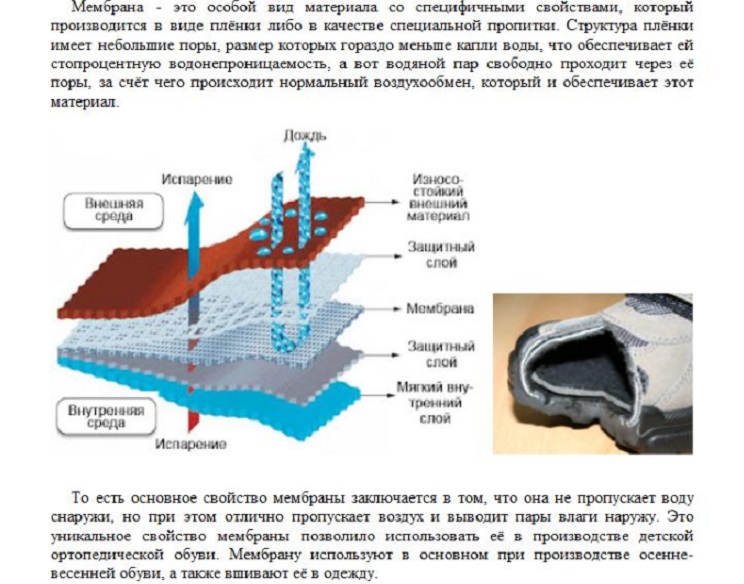 Материал применяется для кровель, подверженных значительным тепловым нагрузкам.
Материал применяется для кровель, подверженных значительным тепловым нагрузкам.
Применение MASTERMAX METAL предоставляет идеальные условия для организации эффективной вентиляции пространства под кровлей. Пленка способна обеспечить непрерывную циркуляцию потоков воздуха, в результате чего влажный воздух, конденсат и пар максимально быстро отводятся из чердачного помещения. Более того, использование этой супердиффузионной мембранной пленки позволяет создать между крышей и утеплителем определенное термическое пространство, благодаря которому в летнее время года уменьшается нагрев дома, в зимний период на поверхности крыши не будет появляться наледь.
Таким образом, в настоящее время на рынке строительных материалов присутствует множество самых разнообразных супердиффузионных мембран, обладающих различными характеристиками. Выбор того или иного типа зависит от конкретных условий эксплуатации и климатической зоны, в пределах которой расположено строение.
pleves24.lv
pleves24.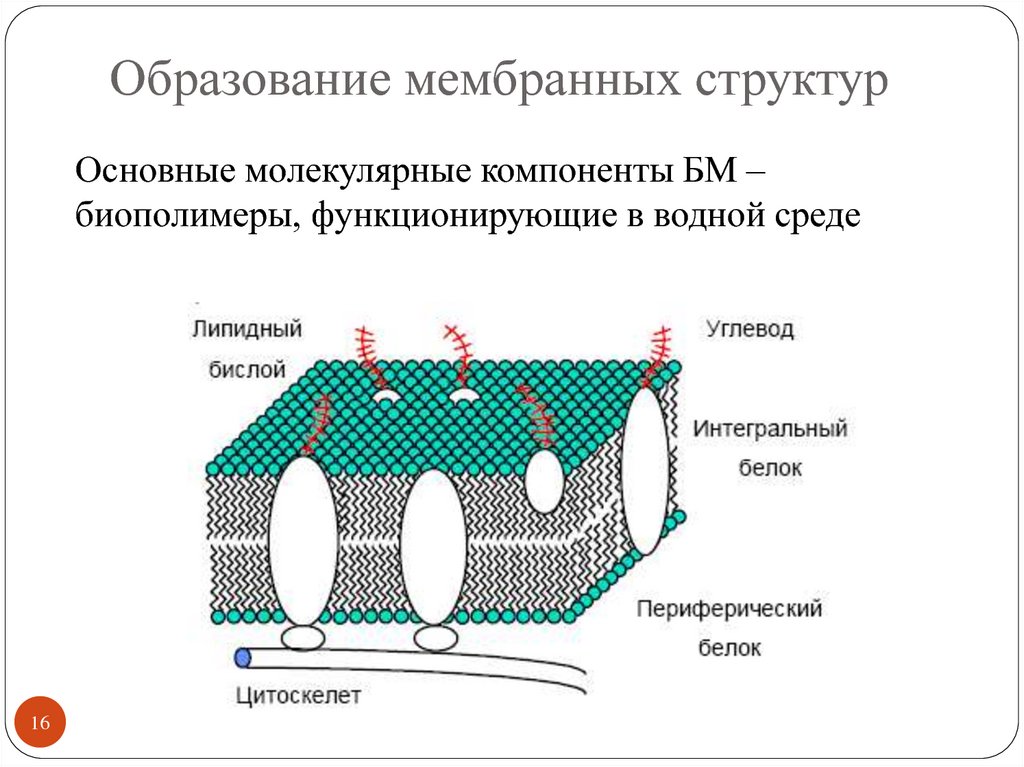 lv
lv
Информация
Как использовать?
Aссортимент
| Заголовок | Описание | Цена | |
| TYVEK Soft |
Высокотехнологичный мембранный материал на основе полиэтилена, представляет собой легкий, чрезвычайно прочный, экологически чистый нетканый материал. Эффективно… |
87.00 € (1.16€/м²) |
ДАЛЕЕ |
| ROOF 100 |
Диффузионная мембрана.Применяется как гидро и ветрозащитный материал с высокой паропроницаемостью для утепленных скатных крыш и стен.Благодаря… |
56.25 € (0.75€/м²) |
ДАЛЕЕ |
| ROOF 130 |
Диффузионная мембрана. |
47.25 € (0.63€/м²) |
ДАЛЕЕ |
| ROOF 150 |
Диффузионная мембрана.Применяется как гидро и ветрозащитный материал с высокой паропроницаемостью для утепленных скатных крыш и стен.Благодаря… |
51.00 € (0.68€/м²) |
ДАЛЕЕ |
| ELKATEK SD 130 + Клейкая лента |
Диффузионная мембрана.Применяется как гидро и ветрозащитный материал с высокой паропроницаемостью для утепленных скатных крыш и стен.Благодаря… |
47.25 € (0.63€/м²) |
ДАЛЕЕ |
| Fortex Opti |
Trīsslāņu elpojoša membrāna sienām un jumtam. |
56.25 € (0.75€/м²) |
ДАЛЕЕ |
| Fortex Extra + 2Tape |
63.75 € (0.85€/м²) |
ДАЛЕЕ | |
| FORTEX Max + Double tape |
|
18.75 € (1.25€/м²) |
ДАЛЕЕ |
| JUTADACH 95 |
Супердиффузионная мембрана.Применяется как гидро и ветрозащитный материал с высокой паропроницаемостью для утепленных скатных крыш и стен.Благодаря… |
63.75 € (0.85€/м²) |
ДАЛЕЕ |
| TYVEK Max Protect |
Основным отличием этой мембраны от Soft и Solid является ее большая прочность на разрыв и высокая. |
178.13 € (2.38€/м²) |
ДАЛЕЕ |
| Tyvek Metal |
Дифузионная мембрана с полипропиленовой сеткой, создающей вентиляционный зазор. Для медных, титановых, металических крыш сложной конструкции. Создает… |
70.13 € (2.13€/м²) |
ДАЛЕЕ |
| JUTADACH 115 |
Супердиффузионная мембрана.Применяется как гидро и ветрозащитный материал с высокой паропроницаемостью для утепленных скатных крыш и стен.Благодаря… |
72.19 € (0.96€/м²) |
ДАЛЕЕ |
| JUTADACH 132 ECO (3 m.) (JUTAECO 2) |
Супердиффузионная мембрана. |
46.50 € (0.62€/м²) |
ДАЛЕЕ |
| JUTADACH Super Plus Double Tape |
Диффузионная мембрана с увеличенной механической прочностью. Применяется как гидро и ветрозащитный материал с высокой паропроницаемостью для… |
193.13 € (2.58€/м²) |
ДАЛЕЕ |
| JUTADREN |
Дифузионная мембрана с полипропиленовой сеткой, создающей вентиляционный зазор. Для медных, титановых, металических крыш сложной конструкции. Создает… |
463.13 € (6.18€/м²) |
ДАЛЕЕ |
| TYVEK Solid |
Высокотехнологичный мембранный материал на основе полиэтилена, представляет собой легкий, чрезвычайно прочный, экологически чистый нетканый материал. |
108.00 € (1.44€/м²) |
ДАЛЕЕ |
| Tyvek SUPRO Plus Tape |
Мембрана c клеящей лентой повышенной прочности с высокой паропроницаемостью, используется для гидроизоляции и ветрозащиты скатных кровель,… |
129.75 € (1.73€/м²) |
ДАЛЕЕ |
| PRIMADACH 100 |
Диффузионная мембрана, пригодна для укладки прямо на утеплитель Применение: для металлических, битумных, шиферных, черепичных, камышовых скатных… |
33.75 € (0.45€/м²) |
ДАЛЕЕ |
| DELTA-VENT |
Оптимальный выбор для надежного решения Качественная трехслойная диффузионная мембрана, пригодна для укладки как прямо на утеплитель,. |
72.00 € (0.96€/м²) |
ДАЛЕЕ |
| DELTA-VENT N |
Идеальное сочетание цены и качества Качественная трехслойная диффузионная мембрана, соответствующая немецким строительным нормам. Слои соединены ультрaзвуком… |
412.50 € (1.38€/м²) |
ДАЛЕЕ |
| DELTA-MAXX |
Позволяет удерживать и постепенно вывести наружу до 1000 г/м2 конденсата. Соответствует классу пожаростойкости b1 (din 4102)…. |
152.25 € (2.03€/м²) |
ДАЛЕЕ |
| PRIMADACH 135 |
Трёхслойная диффузионная мембрана, пригодна для укладки прямо на утеплитель |
48. |
ДАЛЕЕ |
| DELTA-MAXX PLUS |
На 9% уменьшает расходы на отопление среднего семейного дома Качественная антиконденсатная диффузионная мембрана с самоклеющей лентой… |
215.63 € (2.88€/м²) |
ДАЛЕЕ |
| DELTA-MAXX TITAN |
Отражает до 50% внешней теплоэнергии Качественная антиконденсатная диффузионная мембрана с отражающим слоем и самоклеющей лентой вдоль… |
121.88 € (1.63€/м²) |
ДАЛЕЕ |
| DELTA TRELA |
Снижает шум падающего дождя и града на 15 дБ. В комплекте каждого рулона — специальные гвозди…. |
– |
ДАЛЕЕ |
| DELTA-FOXX |
Не имеет аналогов. |
– |
ДАЛЕЕ |
| DELTA-GEO-DRAIN Quattro |
Надежный дренаж и защита окружающей среды Четырёхслойная геомембрана для обустройства дренажа в условиях усилённого гидростатического давления |
– |
ДАЛЕЕ |
 Применяется как гидро и ветрозащитный материал с высокой паропроницаемостью для утепленных скатных крыш и стен.Благодаря…
Применяется как гидро и ветрозащитный материал с высокой паропроницаемостью для утепленных скатных крыш и стен.Благодаря… Plēvei ir elastīgs funkcionāls slānis, kas ir pilnīgi necaurlaidīgs pret…
Plēvei ir elastīgs funkcionāls slānis, kas ir pilnīgi necaurlaidīgs pret… ..
.. Применяется как гидро и ветрозащитный материал с высокой паропроницаемостью для утепленных скатных крыш и стен.Благодаря…
Применяется как гидро и ветрозащитный материал с высокой паропроницаемостью для утепленных скатных крыш и стен.Благодаря… Эффективно…
Эффективно… ..
.. 75 € (0.65€/)
75 € (0.65€/)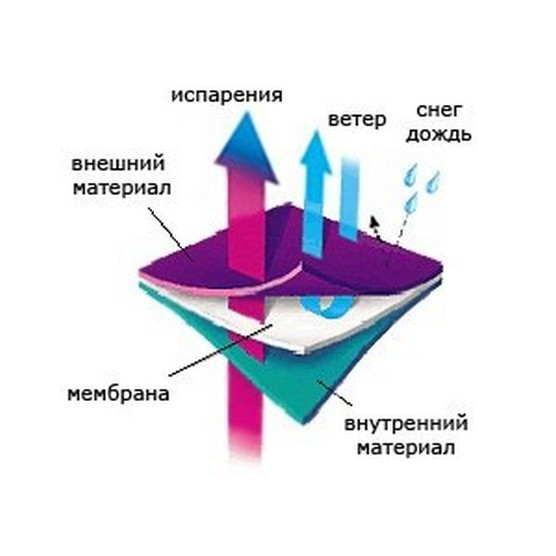 Укладывается только на сплошной настил. Для кровли с малым углом наклона (10–25 градусов)…
Укладывается только на сплошной настил. Для кровли с малым углом наклона (10–25 градусов)…Этот веб-сайт использует файлы cookie — небольшие текстовые файлы, сохраняющиеся на вашем компьютере или мобильном устройстве при открытии веб-сайта. Узнать больше!
Гидроизоляция диффузная мембрана
Главная » Разное » Гидроизоляция диффузная мембрана
Диффузионная мембрана (пленка): Что это такое
Диффузионная мембрана — это рулонный материал для проведения гидроизоляции. Его используют для наружных работ и он является важным звеном в утеплении дома. Материал характеризуется односторонней проводимостью влаги и способен удалять пар из утеплителя, который образуется из-за перепадов температур и существенного повышения влажности.
Его используют для наружных работ и он является важным звеном в утеплении дома. Материал характеризуется односторонней проводимостью влаги и способен удалять пар из утеплителя, который образуется из-за перепадов температур и существенного повышения влажности.
Строение диффузионной мембраны
Она обычно трехслойная, верхний и нижний слой — это полотна из нетканого полипропилена. В мембранах Ондутис средний слой устойчив к растяжениям. Вместе с крепким двухслойным каркасом мембрана характеризуется высокой стойкостью к механическим повреждениям, гидрофобностью и паропроницаемостью. Слои в диффузионных мембранах соединяются с помощью ультразвука — это исключает возможность повреждения середины и позволяет достичь отличных параметров. Такие мембраны экологически безопасны и не подвержены пагубному воздействию бактерий.
Паропроницаемость — важный параметр диффузионной мембраны. По этой характеристике материал делится на три категории:
- Мембраны малой диффузии материал, паропроницаемость которого меньше 300 мг пара на 1 м2 за сутки.
 Такое полотно применяется во внутренних перегородках.
Такое полотно применяется во внутренних перегородках. - Материал средней диффузии проводит от 300 до 1000 мг/м2 за сутки. Обычно применяют именно этот тип мембран, они хорошо зарекомендовали себя в зонах среднего и умеренного климата.
- Мембраны высокой диффузии или супердиффузионные имеют паропроницаемость более 1000 мг/м2/сутки. Они применяются в районах с большими перепадами влажности, суровым климатом или при большой толщине утеплителя.
Для чего предназначена диффузионная мембрана
Диффузионные мембраны защищают утеплитель, внутренние элементы конструкции кровли, стен и всего дома от потерь тепла. Водоотталкивающие свойства материала создают хорошую защиту от внешних воздействий, а воздухонепроницаемость позволяет уменьшить потери тепла в зимний период.
Мембрана защищает утеплитель от негативного воздействия влаги, которая проникает извне под наружную облицовку. Высокая паропроницаемость быстро удаляет воду из-под кровельного пирога.
Мембрана проводит влагу только в одностороннем направлении — от утеплителя, затем она стекает или испаряется. Стороны мембраны маркируются разными цветами, с нанесением логотипа или рисунка на одной из сторон. Ошибочный монтаж материала противоположной стороной обязательно приведет к намоканию теплоизолятора и нарушению утепления дома.
Где используется диффузионная мембрана
Область применения мембран довольно широкая: они могут быть использованы для гидроизоляции крыш, у которых есть подкровельная вентиляция, для теплоизоляционной защиты стропил по всей их высоте, в вентиляционных навесных фасадах и в несущих конструкциях из дерева. При использовании такой мембраны, выполнять дополнительную пропитку дерева необязательно.
Также диффузионная мембрана используется в вентилируемых помещениях и скатных кровлях, не имеющих утепления, где кровельным материалом является:
- полимерная черепица;
- шифер;
- керамическая черепица;
- ондулин;
- металлочерепица.

Преимущества диффузионных мембран
- надежность и высокая прочность материала;
- стойкость к перепадам температуры. Даже сильный перегрев мембраны не приводит к образованию вредных веществ;
- химическая стойкость к кислотам и щелочам;
- безопасность для здоровья.
Особенности монтажа диффузионной мембраны
Важно смонтировать мембрану нужной стороной (в случае с Ондутис — логотипом наружу, а чистой стороной к утеплителю). При монтаже диффузионных мембран важно исключить попадание влаги на теплоизоляцию через стыки и неплотности. Также обязательно нужно изолировать места, где пересекаются проникающие элементы конструкции: вентиляционные короба, антенны, каминные и печные трубы. Там, где они пересекаются, в мембране делается надрез в форме трапеции.
Чтобы мембрана эффективно функционировала, стоит предусмотреть естественную изоляцию пространства под кровлей и обеспечить удаление водяного пара. За эту функцию отвечает вентилируемый зазор между пленкой и покрытием. Его толщина равна толщине обрешетки и контррейки (обычно 7-12 см). Чтобы обеспечить доступ свежему воздуху и удалить водяной пар, делают отверстия для вентиляции.
За эту функцию отвечает вентилируемый зазор между пленкой и покрытием. Его толщина равна толщине обрешетки и контррейки (обычно 7-12 см). Чтобы обеспечить доступ свежему воздуху и удалить водяной пар, делают отверстия для вентиляции.
При наружном утеплении стен диффузионные мембраны монтируются вплотную к внешней стороне теплоизоляции. Материалы крепятся на несущий каркас с помощью строительного степлера или оцинкованных гвоздей. Сверху монтируются контррейки для создания вентиляционного зазора между внутренней стороной облицовочного материала и поверхностью пленки.
Что такое супердиффузионная мембрана
Супердиффузионная мембрана — материал нового поколения. Она будет незаменимой в области с суровым климатом и высокой влажностью, предназначена для комплексной защиты теплоизоляции и отличается от обычной мембраны высокой паропроницаемостью (более 1000 мг/м2/24 часа).
Cупердиффузионные мембраны Ондутис SA115 и SA130 пропускают водяной пар, но задерживают воду и воздух, защищают утеплитель и внутренние элементы стен и кровли. Они сохраняют утеплитель, уменьшают потери тепла кровельного пирога и могут использоваться как временная кровля до монтажа окончательной крыши в течение двух месяцев.
Они сохраняют утеплитель, уменьшают потери тепла кровельного пирога и могут использоваться как временная кровля до монтажа окончательной крыши в течение двух месяцев.
10 голосов , пожалуйста, оцените статью:
Жидкая гидроизоляционная мембрана для гидроизоляции бетонных конструкций
Жидкая гидроизоляционная мембрана доступна в виде жидкости и распыляется или наносится кистью или валиком на бетонную поверхность, которая образует толстую мембрану без швов при контакте с воздухом.
Преимущества жидкой гидроизоляционной мембраны по сравнению с другими традиционными методами Преимущества этого процесса заключаются в его простоте применения.
Процесс без стыков является наиболее важным преимуществом, так как практически отсутствует вероятность просачивания воды , в отличие от WPM рулонного листа и других методов гидроизоляции.Подрядчик может покрыть большую площадь за день и, если нанести на следующий день, может начать перекрытие слоя, законченного в последний день, с помощью гидроизоляционной мембраны с распыляемой жидкостью.
Меры предосторожности:
Образованная таким образом мембрана должна быть подходящей толщины, так как более тонкая мембрана может сломаться, поэтому установщик и ответственный инженер должны соблюдать осторожность.
Характеристики жидкой гидроизоляционной мембраныЭти системы жидкого нанесения характеризуются покрытиями, обладающими свойствами удлинения, долговечностью, гибкостью, абразивной стойкостью и химической стойкостью, что обеспечивает успешную установку.Жидкая гидроизоляционная мембрана обеспечивает оптимальную производительность и, что самое главное, долговечность.
Таким образом, этот метод гидроизоляции экономит время, а также не требует прекращения других работ, так как нанесение утром становится сухим и стойким до полудня. Это значительно экономит трудозатраты и дает максимально качественные результаты.
Хотя листы и другие мембраны со временем разрушаются, нанесенная жидкая гидроизоляция обеспечивает длительный эффект. При нанесении на бетонную поверхность заполняет трещины в бетоне, создавая защитную систему пароизоляции, проникает в бетон на глубину до 20 мм и сохраняет водонепроницаемость, тем самым увеличивая прочность бетона.
При нанесении на бетонную поверхность заполняет трещины в бетоне, создавая защитную систему пароизоляции, проникает в бетон на глубину до 20 мм и сохраняет водонепроницаемость, тем самым увеличивая прочность бетона.
Во многих странах, таких как США, Великобритания, эту гидроизоляционную мембрану наносят также на наружные стены. Он доступен в разных цветах, чтобы он соответствовал цвету стены. Прибл. 5 галлонов гидроизоляционной мембраны можно использовать на 325 кв.м бетонной поверхности. Вам нужно нанести 1 слой, дать ему высохнуть, а затем нанести второй слой.
Может также применяться на бетонных мостах (может иметь норму нанесения 800 кв.метр в час). Бетон расширяется и сжимается, вызывая трещины при высоких или низких температурах, вызывая просачивание воды, жидкая гидроизоляционная мембрана обладает свойствами поглощения ультрафиолета и, таким образом, защищает бетон от повреждений в таких условиях.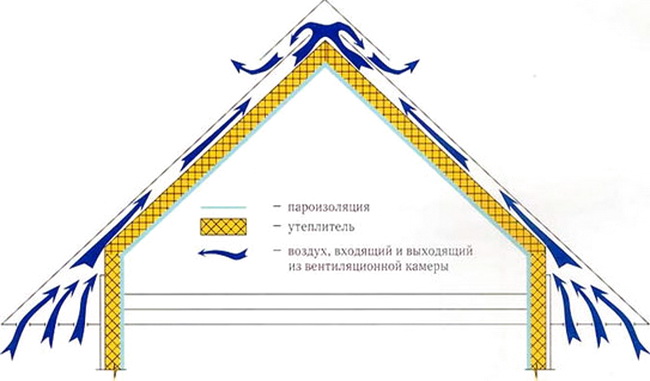
Жидкую гидроизоляционную мембрану можно также наносить на стальные мосты с металлическим настилом, которые имеют систему стыков космической эры. Это предотвращает просачивание из суставов. Это увеличивает долговечность моста, и это становится очевидным, когда мост восстанавливается через 10-15 лет.
Жидкая гидроизоляционная мембрана не только предотвращает образование трещин, но и предотвращает коррозию внутренних материалов, стальных стержней и стыков. Он герметизирует трещины шириной до 1,3 мм и выдерживает расширение до 0,4 мм, и более того, мост готов к эксплуатации менее чем за 6 часов.
Подготовка поверхности под нанесение жидкой гидроизоляционной мембраныГидроизоляционная поверхность должна быть чистой от грязи, сухой и чистой.На поверхности не должно быть свободных пятен, поэтому подрядчик должен обеспечить надлежащую подготовку поверхности перед нанесением.
Отверждение жидкой гидроизоляционной мембраны Очень важно поливать нанесенную мембрану сразу после высыхания мембраны на ощупь. Согласно стандартным спецификациям, через пару дней его необходимо вылечить водой.
Согласно стандартным спецификациям, через пару дней его необходимо вылечить водой.
Обеспечивает низкие эксплуатационные расходы и долгий срок службы. Это разумная альтернатива на всю жизнь другим традиционным методам.Если мост прослужит 200 лет, значит, гидроизоляция тоже работает.
Это также позволяет сэкономить на наполнителях и герметиках и других материалах, которые будут использоваться в будущем для ремонта трещин, и по-прежнему остается только временным решением.
Подробнее о методах гидроизоляции
.
Жидкая гидроизоляционная мембрана | GCP Applied Technologies
Жидкая гидроизоляция имеет множество преимуществ, в том числе:
- Простота нанесения – Жидкую гидроизоляцию часто можно нанести быстро. В зависимости от продукта бригада из трех человек может нанести распылением 10 000 квадратных футов или 930 квадратных метров гидроизоляции всего за один день.
 Эти преимущества становятся еще более заметными при гидроизоляции сложных пространств или пространств с большим количеством деталей, поскольку гидроизоляцию труб и других проходов с помощью жидкости легче, чем путем разрезания и размещения листовых мембран.
Эти преимущества становятся еще более заметными при гидроизоляции сложных пространств или пространств с большим количеством деталей, поскольку гидроизоляцию труб и других проходов с помощью жидкости легче, чем путем разрезания и размещения листовых мембран. - Бесшовные – Гидроизоляция, наносимая жидкостью, затвердевает, образуя одну сплошную мембрану, поэтому отсутствуют швы или стыки, которые являются наиболее частыми областями, где гидроизоляция имеет тенденцию к разрушению.
- Гибкость – Самые эффективные системы жидкой гидроизоляции обладают гибкостью, чтобы расширяться и сжиматься при изменении температуры и адаптироваться к незначительным структурным сдвигам. Таким образом, гидроизоляция продолжает работать даже при экстремальных погодных условиях и / или при появлении небольших трещин в бетоне в основной конструкции.
Применение жидкой гидроизоляционной мембраны
Жидкая гидроизоляционная мембрана обычно используется в самых разных областях, например:
- Зеленые крыши – Жидкие гидроизоляционные мембраны хорошо подходят для защиты зеленых крыш.
 Обычно зеленые крыши имеют много проходов для полива. На детализацию несложно нанести жидкую гидроизоляцию. Убедитесь, что выбранная вами система гидроизоляции совместима с конструкцией зеленой крыши и проверена в аналогичных условиях.
Обычно зеленые крыши имеют много проходов для полива. На детализацию несложно нанести жидкую гидроизоляцию. Убедитесь, что выбранная вами система гидроизоляции совместима с конструкцией зеленой крыши и проверена в аналогичных условиях.Если вы планируете проект зеленой крыши, убедитесь, что выбранная вами гидроизоляционная система выдерживает длительные испытания на устойчивость к корням, чтобы не повредить ее при укоренении растений.
- Скрытые крыши – Скрытая крыша часто представляет собой часть подвала, которая выходит за пределы основного возвышения и обычно является частью подсадной конструкции или ландшафтной зоны. Подъемный настил, используемый людьми или автомобилями, также можно отнести к категории заглубленных конструкций крыши.
Гидроизоляционные системы для подземных крыш должны служить в течение всего срока службы конструкции, потому что ремонт гидроизоляционной системы разрушителен для владельцев и жителей здания.
Ищите гидроизоляционную систему, которая является чрезвычайно прочной, а также предлагает гибкость для адаптации к изменениям температуры и другим факторам, которые могут повредить некоторые гидроизоляционные мембраны.
 Плотное соединение между гидроизоляционной системой и основанием также имеет решающее значение для предотвращения миграции воды под поверхность.
Плотное соединение между гидроизоляционной системой и основанием также имеет решающее значение для предотвращения миграции воды под поверхность. - Перевернутая крыша – В некоторых проектах, таких как террасы на крыше, используется конструкция перевернутой крыши. В этих сценариях гидроизоляционный слой устанавливается под традиционной изоляцией крыши. Утеплитель кладется поверх жидкой гидроизоляционной мембраны и закрепляется тротуарной плиткой или щебнем. Бесшовная природа жидкой гидроизоляции делает ее подходящей для этих проектов.Жидкость легко наносится. После затвердевания образует одну прочную мембрану, устойчивую к утечкам.
В прошлом генеральные подрядчики полагались на нанесение горячей гидроизоляции из-за ее низкой стоимости и способности хорошо прилегать к основанию. В зависимости от региона генеральные подрядчики могут столкнуться с дополнительными разрешениями, должны будут доплачивать за работу пожарных на объекте и / или столкнуться с препятствиями, обеспечивающими страхование ответственности при использовании горячей гидроизоляции.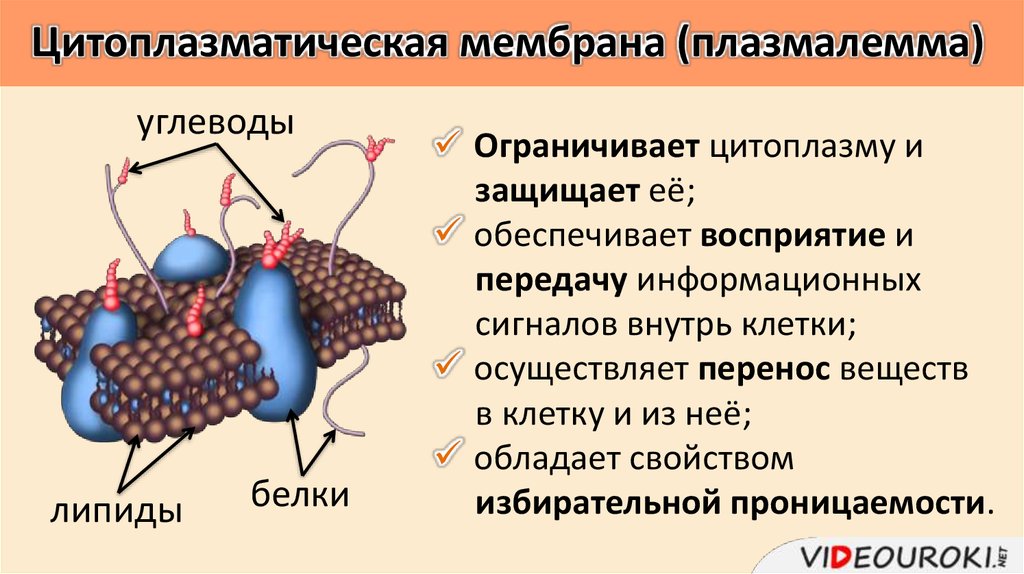
Для многих генеральных подрядчиков дополнительные проблемы означают, что нанесение горячей гидроизоляции больше не имеет смысла, особенно когда доступны варианты холодной гидроизоляции, которые предлагают такие же или лучшие адгезионные свойства.Использование холодной гидроизоляции снижает сложность и риски во время строительства и ускоряет процесс установки.
Связанные
.
9235 Гидроизоляционная мембрана | Интерьер и экстерьер
9235 Гидроизоляционная мембрана – это простая в установке тонкая несущая гидроизоляционная мембрана, разработанная специально для особых требований при укладке керамической плитки, камня и кирпича. Это идеальное решение для влажных помещений и при длительном погружении в воду.
9235 Гидроизоляционная мембрана представляет собой тонкую (0,02 дюйма [0,5 мм]) несущую, гибкую, наносимую жидкостью гидроизоляционную мембранную систему, состоящую из жидкого каучукового полимера, который наносится холодным способом с интегрированной армирующей тканью, чтобы обеспечить бесшовную водонепроницаемую мембрану. Его быстро наносят кистью или валиком с длинным ворсом, чтобы сформировать гибкую бесшовную гидроизоляционную мембрану, которая сцепляется с широким спектром субстратов (включая как обычные, так и менее обычные коммерческие субстраты). 9235 Гидроизоляционная мембрана также содержит антимикробную защиту для предотвращения роста плесени и грибка.
Его быстро наносят кистью или валиком с длинным ворсом, чтобы сформировать гибкую бесшовную гидроизоляционную мембрану, которая сцепляется с широким спектром субстратов (включая как обычные, так и менее обычные коммерческие субстраты). 9235 Гидроизоляционная мембрана также содержит антимикробную защиту для предотвращения роста плесени и грибка.
Оснащен антимикробной защитой Microban
.
Диффузионные мембраны: использование, монтаж
В монтаже строительных конструкций помимо базовых свойств и эксплуатационных качеств используемых материалов важную роль играют вспомогательные элементы. В их числе особое место занимают изоляторы, обеспечивающие целостность основной структуры. Существенна функция данного компонента в кровельных покрытиях. Благодаря изоляционной прослойке формируется барьер перед осадками и другими внешними воздействиями. На рынке представлены разные по характеристикам средства обеспечения такой защиты, но самым эффективным материалом являются гидроизоляционные диффузионные мембраны, которые закладываются непосредственно в конструкцию крыши. Но кровельными системами не ограничиваются сферы применения данного покрытия. Например, изолятор также используют в укладке напольных и настенных материалов.
В их числе особое место занимают изоляторы, обеспечивающие целостность основной структуры. Существенна функция данного компонента в кровельных покрытиях. Благодаря изоляционной прослойке формируется барьер перед осадками и другими внешними воздействиями. На рынке представлены разные по характеристикам средства обеспечения такой защиты, но самым эффективным материалом являются гидроизоляционные диффузионные мембраны, которые закладываются непосредственно в конструкцию крыши. Но кровельными системами не ограничиваются сферы применения данного покрытия. Например, изолятор также используют в укладке напольных и настенных материалов.
Общие сведения о диффузных изоляторах
Специально для сохранения технико-физических свойств настила кровли строители используют гидроизоляционные подкровельные пленки. Большинство из них реализуется в виде рулонов, которые внешне практически ничем не отличаются друг от друга. Оценивать же такие изоляторы можно по разным критериям, среди которых паропроницаемость. Собственно, это основное качество полимерных пленок, которое обуславливает возможность их структуры пропускать водяной пар. На языке специалистов этот параметр называется плотностью диффузионного потока. На практике использования паропроницаемость обуславливает интенсивность прохождения влаги через крышу в используемые помещения. Таким образом, диффузионные мембраны минимизируют возможность проникновения осадков и их производных под кровлю. Аналогичные функции выполняют практически все специализированные гидроизоляторы, но к мембранам относят и способность противостоять интенсивным воздействиям осадков в условиях порывистого ветра. Также если обычные изоляционные прослойки формируют лишь базовую защиту от дождевых капель и снега, то диффузионная пленка становится эффективной преградой перед водяным паром.
Собственно, это основное качество полимерных пленок, которое обуславливает возможность их структуры пропускать водяной пар. На языке специалистов этот параметр называется плотностью диффузионного потока. На практике использования паропроницаемость обуславливает интенсивность прохождения влаги через крышу в используемые помещения. Таким образом, диффузионные мембраны минимизируют возможность проникновения осадков и их производных под кровлю. Аналогичные функции выполняют практически все специализированные гидроизоляторы, но к мембранам относят и способность противостоять интенсивным воздействиям осадков в условиях порывистого ветра. Также если обычные изоляционные прослойки формируют лишь базовую защиту от дождевых капель и снега, то диффузионная пленка становится эффективной преградой перед водяным паром.
Принцип действия мембраны
Распространенной проблемой многих гидроизоляторов является скопление влаги в одном месте – как правило, перед непроницаемым слоем. Грамотный монтаж предусматривает специальные каналы отвода жидкости, но и этот способ не избавляет от всех проблем эксплуатации. По другому принципу работают диффузионные мембраны, включенные в состав кровельных «пирогов». Он действует не пассивно, а активно, то есть не обеспечивают стационарный барьер, а за счет специального слоя проводит влагу от утеплителя к месту будущего стекания или выветривания. Поэтому очень важно правильной стороной располагать материал в рабочей зоне.
По другому принципу работают диффузионные мембраны, включенные в состав кровельных «пирогов». Он действует не пассивно, а активно, то есть не обеспечивают стационарный барьер, а за счет специального слоя проводит влагу от утеплителя к месту будущего стекания или выветривания. Поэтому очень важно правильной стороной располагать материал в рабочей зоне.
Сама структура функционального полотна таких изоляторов напоминает капиллярный насос, который формируется двумя прослойками. Ворсистая ткань выполняет функцию сборщика влаги и передает ее к участкам с перфорацией. Далее по микроскопическим отверстиям капли с паром просачиваются на внешнюю сторону полотна. В такой схеме влагоотвода есть риск контакта жидкости с металлическими элементами кровельной конструкции. Чтобы предотвратить процессы коррозии, используется объемная диффузионная мембрана на полипропиленовой нетканой основе. Также сберечь восприимчивые к влажности материалы помогут пленки с повышенной пропускной способностью – они быстрее обеспечивают вывод влаги, сокращая шансы возникновения разрушительных процессов.
Разновидности мембран
Различаются покрытия этого типа по нескольким критериям, но главным считается степень паропроницаемости. В частности, выделяют низко- и высокопаропроницаемые изоляторы. В первую категорию входят диффузионные мембраны, которые состоят из 2-3 слоев с армирующей сеткой. Такой материал обладает средними способностями к проведению водяного пара, но как универсальное средство защиты от осадков может стать оптимальным решением. Высокопаропроницаемые материалы демонстрируют более высокий уровень пропускной способности, из-за чего их также называют супердиффузионными пленками. Опять же, высокая скорость проведения влаги не позволяет ей конденсироваться, что способствует предохранению утепляющего слоя от разрушения. По этой причине строители упрощают изоляционные прослойки, избавляя их от вентиляционных зазоров.
Также в некоторых случаях используются специальные виды паропроницаемых пленок, среди которых и упомянутые объемные мембраны. Этот материал обычно применяют для обеспечения изоляции фальцевых и металлических покрытий. Монтируют объемные разделительные диффузионные мембраны с обеспечением верхнего вентиляционного зазора на кровлях, изготовленных из алюминия, стали, титана и других металлов.
Монтируют объемные разделительные диффузионные мембраны с обеспечением верхнего вентиляционного зазора на кровлях, изготовленных из алюминия, стали, титана и других металлов.
Характеристики диффузных мембран
Основным эксплуатационным показателем таких материалов является паронепроницаемость. Но при выборе также учитываются и другие характеристики – например, грамматура, водонепроницаемость и температурная стойкость. Что касается грамматуры, то она обозначает массу пленки, которая увеличивается пропорционально толщине материала. Также по мере увеличения веса повышается и механическая прочность изоляции. На рынке можно найти пленки, грамматура которых варьируется от 60 до 270 г/м2. Также имеет значение и устойчивость покрытия к водяному давлению. Этой характеристикой обозначается водонепроницаемость, которой обладает диффузионная мембрана в конкретном исполнении. Существуют разные методики оценки этого параметра, но самой простой считается система, предусматривающая маркировку по классам W1-W3. Так, принадлежность мембраны к категории W1 указывает на то, что материал в течение 2 часов сможет выдерживать давление водяного столба, высота которого составляет 20 см.
Так, принадлежность мембраны к категории W1 указывает на то, что материал в течение 2 часов сможет выдерживать давление водяного столба, высота которого составляет 20 см.
Кроме осадков и влажности, вред гидроизоляционным пленкам способна нанести теплая солнечная погода. В стандартном исполнении такие материалы выдерживают пиковое температурное воздействие в диапазоне 80-100 °C. Что касается стойкости к ультрафиолету, то, например, диффузионная мембрана «Технониколь» в базовой версии может находиться под прямыми лучами солнца 3 месяца. Для усиленных модификаций этот временной промежуток увеличивается до 4 месяцев.
Применение в кровельной конструкции
В составе кровли изоляционная мембрана обеспечивает защиту утеплителя и внутренних частей конструкции. Прослойка выполняет несколько функций одновременно – кроме предохранения от влаги и пара она также ограждает элементы крыши от выветривания. Неправильно относить задачи данного материала к изоляции помещений здания.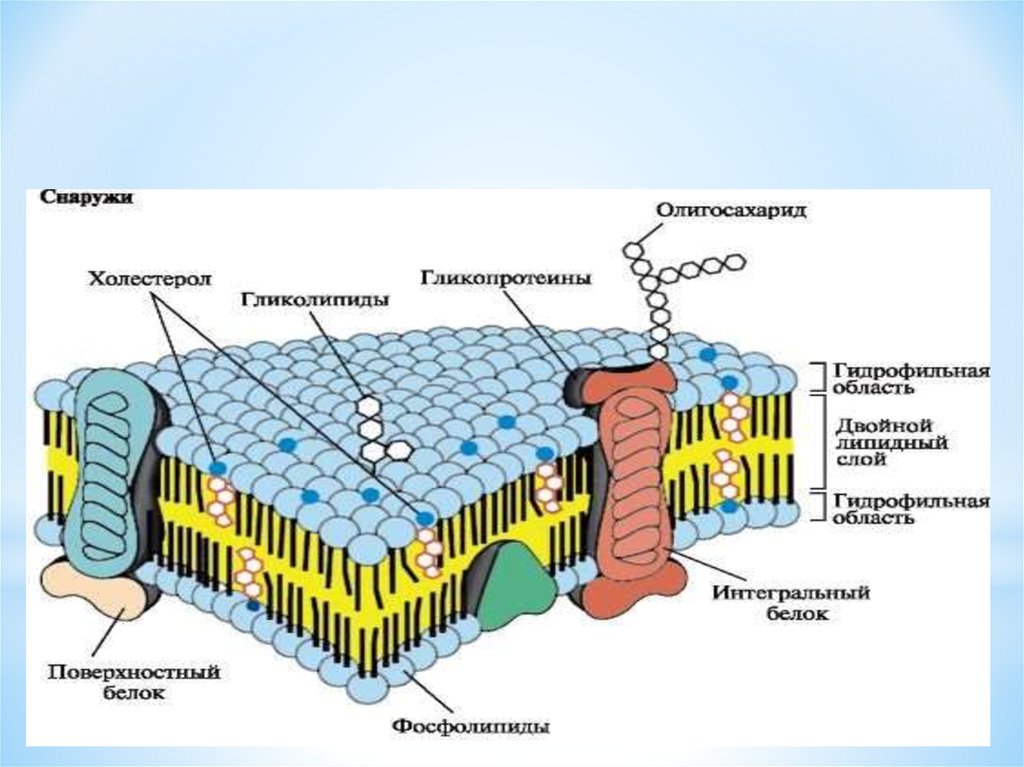 Основные функции мембраны все же распространяются на защиту утепляющего слоя, который, в свою очередь, регулирует влажностный, а в некоторых случаях и температурный режимы. В ходе проектирования схемы устройства мембраны необходимо предотвратить возможности стекания воды с пленочной поверхности на теплоизолятор по местам стыков. Как правило, подкровельная диффузионная мембрана соседствует с каналами вентиляции, каминными и печными трубами, стойками антенн и другими сторонними компонентами, пересечения с которыми необходимо изолировать.
Основные функции мембраны все же распространяются на защиту утепляющего слоя, который, в свою очередь, регулирует влажностный, а в некоторых случаях и температурный режимы. В ходе проектирования схемы устройства мембраны необходимо предотвратить возможности стекания воды с пленочной поверхности на теплоизолятор по местам стыков. Как правило, подкровельная диффузионная мембрана соседствует с каналами вентиляции, каминными и печными трубами, стойками антенн и другими сторонними компонентами, пересечения с которыми необходимо изолировать.
Применение мембран в устройстве напольных покрытий
Обеспечение надежной изоляции требуется и в самих помещениях. Так, напольные поверхности сохраняют декоративные и технические качества только в условиях оптимального температурно-влажностного режима. Конечно, есть материалы, которые не чувствительны к тем же воздействиям влаги, но древесные поверхности к таким не относятся. Обычно применение диффузионных мембран в напольных покрытиях ограничивается созданием вспомогательной подложки для утеплителя. Впрочем, может иметь значения этаж, на котором выполняются работы. Самым требовательным является нижний уровень в частном доме, особенно если он связывает жилые помещения с подвалом.
Впрочем, может иметь значения этаж, на котором выполняются работы. Самым требовательным является нижний уровень в частном доме, особенно если он связывает жилые помещения с подвалом.
Применение в облицовке стен и фасадов
С помощью диффузионной пленки также обеспечивают защиту от выветривания и влаги теплоизоляционных материалов, находящихся в составе вентилируемых фасадов. Мембраны размещаются с внешней стороны, обеспечивая, таким образом, возможность прохождения излишков влаги и пара через вентилируемый зазор в облицовке. Установка производится непосредственно на фасаде дома вместе с теплоизоляционными панелями. Кстати, если в конструкции используется металлический несущий каркас, то оптимально подойдет объемная диффузионная мембрана, которая создаст естественный сток влаги без риска развития коррозии. Если планируется использовать в отделке сайдинг или вагонку, то пленку нужно фиксировать вплотную к наружной стороне изоляции под декоративной облицовкой. Это один из простейших способов применения диффузных изоляторов, так как он требует минимальных усилий в плане монтажа – достаточно воспользоваться строительным степлером или оцинкованными гвоздиками и закрепить материал к несущей конструкции.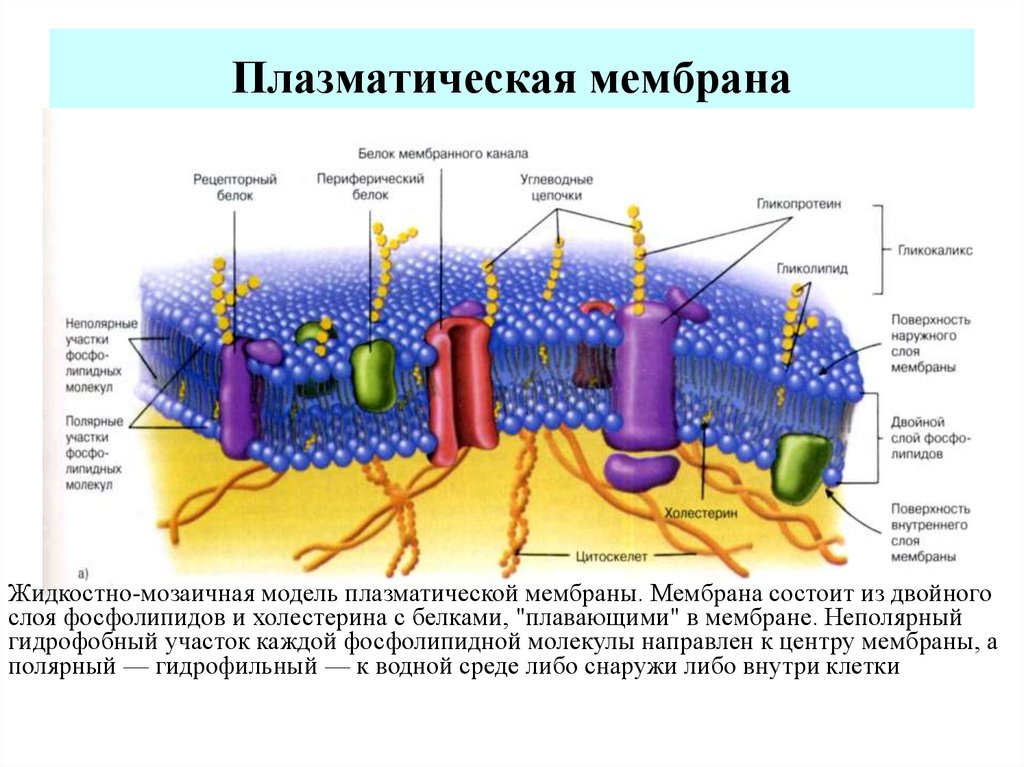
Монтаж диффузионной мембраны
При наружном способе размещения материала желательно выполнять операцию в сухую погоду и вместе с фиксацией теплоизолятора. Мембрану следует раскатать по всей рабочей поверхности и закрепить ее гвоздиками или скобками. Классическая установка на крыше предполагает укладку горизонтальными полосками с нахлестом порядка 15 см. При этом вертикальные стыки окончаний следует размещать на стропилах. Все точки и линии промежутков между отдельными кусками материала фиксируются с помощью бутилкаучуковой или акриловой ленты. Вентиляционный зазор не требуется, если диффузионные мембраны планируется устанавливать на крыше, но при укладке изолятора на фасадных поверхностях его лучше предусмотреть. В зимнее время нагрузка на утепляющий материал значительно повышается, поэтому рекомендуется осенью выполнять ревизию кровли на предмет плотности прилегания мембраны к изолятору. Если у стропил толщина превышает аналогичный показатель теплоизолятора, то паронепроницаемый материал следует прижать к их боковой стороне рейками с типоразмером 2х3 см. Также для выполнения этой задачи можно использовать строительные скобы. В ходе работы следует ориентироваться на то, чтобы нижняя кромка мембраны обеспечивала беспрепятственное удаление влаги в специальный желоб водоотвода.
Также для выполнения этой задачи можно использовать строительные скобы. В ходе работы следует ориентироваться на то, чтобы нижняя кромка мембраны обеспечивала беспрепятственное удаление влаги в специальный желоб водоотвода.
Производители
Нетканый паронепроницаемый материал представляют на рынке разные изготовители, среди которых немало и отечественных. В частности, диффузионная мембрана «Технониколь» позволяет устраивать гидроизоляцию, которая одновременно защищает конструкции от контактов с влагой и дает поверхностям «дышать». Материал этой линейки представлен в нескольких версиях, в том числе и с усиливающими волокнами. Также у пользователей завоевала доверие супердиффузионная пленка «Технониколь», которая выполнена из трехслойного микропористого материала. Отличием данного изолятора является двухстороннее применение нетканого полипропилена, выполняющего защитную функцию для основной прослойки. Такую мембрану целесообразно использовать именно в кровельных покрытиях, которые требуют не только качественного выполнения изоляционных задач, но и сохранения прочностных свойств конструкции.
Довольно известным изготовителем паронепроницаемых гидроизоляторов является и компания Du Pont, специализирующаяся как раз на защите стеновых и кровельных систем посредством воздушной и водной фильтрации. Под этим брендом выходит диффузионная мембрана Tyvek, представляющая собой многофункциональную гидроветрозащитную пленку. Материал имеет сетчатую структуру, образованную полимерными волокнами. В отличие от конкурирующих изоляторов покрытия Tyvek отличаются экологической чистотой. То есть их можно использовать и в наружной отделке, и в структуре внутренних облицовочных покрытий. В обоих случаях включение диффузионной пленки этой марки будет обеспечивать сухость и долговечность отделки.
Заключение
Традиционные способы устройства изоляционных прослоек обычно не требуют соблюдения особых условий монтажа. Достаточно подобрать лист или плиту соответствующих размеров и зафиксировать материал на рабочем участке. По такому принципу обычно монтируется и утепляющий материал, и гидроизоляция. Диффузионная мембрана, в свою очередь, гарантирует более высокий эксплуатационный эффект, чем обычные изоляторы. Но для его достижения важно соблюдать правила расположения и крепления паронепроницаемых пленок относительно утепляющих слоев. Кровельные конструкции в этом отношении являются наиболее сложными, так как выводящий влагу материал может контактировать с другими компонентами покрытия и системами крыши. Также изначально следует предусмотреть коммуникации для водоотвода, с которыми будет взаимодействовать диффузионный материал. С точки зрения установки, несколько проще выглядит монтаж изоляторов на стенах и фасадных конструкциях. В таких случаях все-таки на первое место выходит задача сохранения вентиляционных свойств покрытия, которые и реализуются благодаря включению паронепроницаемой мембраны в состав утепляющей прослойки.
Диффузионная мембрана, в свою очередь, гарантирует более высокий эксплуатационный эффект, чем обычные изоляторы. Но для его достижения важно соблюдать правила расположения и крепления паронепроницаемых пленок относительно утепляющих слоев. Кровельные конструкции в этом отношении являются наиболее сложными, так как выводящий влагу материал может контактировать с другими компонентами покрытия и системами крыши. Также изначально следует предусмотреть коммуникации для водоотвода, с которыми будет взаимодействовать диффузионный материал. С точки зрения установки, несколько проще выглядит монтаж изоляторов на стенах и фасадных конструкциях. В таких случаях все-таки на первое место выходит задача сохранения вентиляционных свойств покрытия, которые и реализуются благодаря включению паронепроницаемой мембраны в состав утепляющей прослойки.
ДИФФУЗИЯ ЧЕРЕЗ КЛЕТОЧНУЮ МЕМБРАНУ
ДИФФУЗИЯ ЧЕРЕЗ КЛЕТОЧНУЮ МЕМБРАНУДИФФУЗИЯ ЧЕРЕЗ КЛЕТОЧНУЮ МЕМБРАНУ
Введение: Вещества, такие как вода, ионы и молекулы, необходимые для клеточных процессов, могут проникать в клетки и покидать их посредством пассивного процесса, такого как диффузия.![]() Диффузия – это случайное движение молекул, но имеющее чистое направление к областям.
более низкой концентрации, чтобы достичь равновесия.
Диффузия – это случайное движение молекул, но имеющее чистое направление к областям.
более низкой концентрации, чтобы достичь равновесия.
Простая пассивная диффузия
происходит, когда небольшие молекулы проходят через липидный бислой клеточной мембраны. Облегченная диффузия зависит от белков-переносчиков, встроенных в мембрану. определенные вещества для прохождения, которые не могут диффундировать через клеточную мембрану.Важность: На скорость диффузии влияют свойства клетки, диффундирующей молекулы и окружающего раствора. Мы можем использовать простые уравнения и графики, чтобы исследовать, как определенные молекулы и их концентрация влияют на скорость диффузии. Мы также можем сравнить простую и облегченную диффузию.
Вопрос: Как различаются скорости простой и облегченной диффузии в зависимости от градиента концентрации?
Простое распространение
Переменные:
п | количество молекул внутри клетки (моль) |
т | время (секунды) |
Р | константа проницаемости для конкретной молекулы (см/с) |
А | площадь поверхности клеточной мембраны (см 2 ) |
С | концентрация диффундирующей молекулы (моль/см 3 ) |
х | ширина клеточной мембраны (см) |
Метод: скорость простой диффузии может быть выражена модификацией закона Фика для небольших неполярных молекул. Скорость диффузии, dn/dt, представляет собой изменение количества диффундирующих молекул внутри клетки с течением времени.
Скорость диффузии, dn/dt, представляет собой изменение количества диффундирующих молекул внутри клетки с течением времени.
Поскольку чистое движение диффундирующих молекул зависит от градиент концентрации, скорость диффузии прямо пропорциональна градиент концентрации (dC/dx) через мембрану. Концентрация градиент, dC/dx, представляет собой разницу в концентрации молекул внутри и вне клетки. через клеточную мембрану шириной dx. Это эквивалентно (C из – C из )/Dx, где C из и C из — концентрации субстрата внутри и снаружи клетки, Dx — ширина клеточной мембраны. Когда концентрация вне клетки (C из ) больше, чем внутри клетки (C из ), концентрация градиент (dC/dx) будет положительным, а чистое движение будет внутри ячейки (положительное значение dn/dt).
Мы можем описать скорость диффузии как прямо пропорциональную градиенту концентрации с помощью следующего уравнения:
, где A — площадь мембраны, а P — константа проницаемости. P представляет собой константу, связывающую легкость проникновения молекулы в клетку в зависимости от размера молекулы и растворимости в липидах.
P представляет собой константу, связывающую легкость проникновения молекулы в клетку в зависимости от размера молекулы и растворимости в липидах.
Обратите внимание, что когда A и P являются константами, это уравнение просто описывает линию, где dn/dt является функцией dC/dx. Если мы построим график зависимости скорости диффузии от градиента концентрации, мы получим простую линейную функцию.
Интерпретация: Обратите внимание, что скорость диффузии увеличивается по мере градиент концентрации увеличивается. Если концентрация молекул вне клетки очень высока по сравнению с концентрацией внутри клетки, скорость диффузии также будет высокой. Если внутренний и внешние концентрации одинаковы (низкий градиент концентрации), скорость диффузии будет низкой.
Облегченное распространение
Переменные:
п | количество молекул внутри клетки |
т | время (секунды) |
В макс. | константа насыщения (моль/см 3 /сек) |
К | константа определения скорости насыщения (моль/см 4 ) |
С | концентрация диффундирующей молекулы (моль/см 3 ) |
х | ширина клеточной мембраны (см) |

Метод: В отличие от простой диффузии облегченная диффузия включает ограниченное количество белков-носителей. При низких концентрациях молекулы проходят через белки-носители аналогично простой диффузии. При высоком растворении
концентрации, однако все белки заняты диффундирующими молекулами. Дальнейшее увеличение концентрации растворенного вещества не изменит скорость диффузии. Другими словами, существует некоторая максимальная скорость диффузии (Vmax), когда все носители про
подростки
насыщены. Поэтому мы не можем использовать простое линейное уравнение для описания скорости диффузии. Скорость диффузии будет увеличиваться с увеличением концентрации растворенного вещества, но должна асимптотически приближаться к скорости насыщения, V максимум . Как быстро
лы
белки-носители становятся насыщенными, можно описать переменной K, градиентом концентрации, при котором скорость диффузии составляет 1/2 Vmax. K и Vmax зависят от свойств диффундирующей молекулы, таких как ее проницаемость (P), а также
площадь поверхности (A) ячейки, но для упрощения мы приводим уравнение как:
Поэтому мы не можем использовать простое линейное уравнение для описания скорости диффузии. Скорость диффузии будет увеличиваться с увеличением концентрации растворенного вещества, но должна асимптотически приближаться к скорости насыщения, V максимум . Как быстро
лы
белки-носители становятся насыщенными, можно описать переменной K, градиентом концентрации, при котором скорость диффузии составляет 1/2 Vmax. K и Vmax зависят от свойств диффундирующей молекулы, таких как ее проницаемость (P), а также
площадь поверхности (A) ячейки, но для упрощения мы приводим уравнение как:
Мы можем построить график зависимости dn/dt от dC/dx, чтобы увидеть, как скорость диффузии изменяется с увеличением концентрации растворенного вещества вне клетки.
Интерпретация: Изобразив это уравнение в виде графика, мы видим, что при низких
концентрации
растворенного вещества скорость диффузии в клетку происходит почти линейно, как и при простой диффузии. Обратите внимание, что при низких концентрациях растворенного вещества наклон намного круче, чем при простой диффузии.
Облегченная диффузия может увеличить скорость диффузии определенных молекул при низких концентрациях. Однако скорость облегченной диффузии выравнивается с увеличением концентрации растворенного вещества. Дополнительное увеличение внешней концентрации растворенного вещества
не может увеличить скорость диффузии после насыщения белков-носителей.
Обратите внимание, что при низких концентрациях растворенного вещества наклон намного круче, чем при простой диффузии.
Облегченная диффузия может увеличить скорость диффузии определенных молекул при низких концентрациях. Однако скорость облегченной диффузии выравнивается с увеличением концентрации растворенного вещества. Дополнительное увеличение внешней концентрации растворенного вещества
не может увеличить скорость диффузии после насыщения белков-носителей.
Вывод: Пассивная диффузия растворенного вещества в клетку линейно связана с концентрацией растворенного вещества вне клетки. Белки-носители увеличивают скорость диффузии, позволяя большему количеству растворенного вещества проникать в клетку. облегченная диффузия, однако приближается к максимальной скорости, когда белки-носители насыщаются растворенным веществом.
Дополнительные вопросы:
Простое распространение
1. Мы построили график зависимости dn/dt от dC/dx. Каков наклон этой линии? Что биологически означает увеличение или уменьшение наклона?
2. Теперь предположим, что градиент концентрации является постоянным. Как изменяется скорость диффузии (dn/dt) в зависимости от площади поверхности (A) клетки и проницаемости (P) диффундирующей молекулы? Постройте график dn/dt как функцию от A или P и опишите
функция.
Теперь предположим, что градиент концентрации является постоянным. Как изменяется скорость диффузии (dn/dt) в зависимости от площади поверхности (A) клетки и проницаемости (P) диффундирующей молекулы? Постройте график dn/dt как функцию от A или P и опишите
функция.
Облегченная диффузия
1. Посмотрите на уравнение облегченной диффузии и найдите горизонтальную асимптоту. Что происходит с dn/dt, когда dC/dx приближается к бесконечности?
2. Попробуйте построить график этого уравнения с различными значениями K. Как это изменит концентрацию, при которой происходит насыщение белков-носителей?
3. Сравните простую и облегченную диффузию глюкозы в эритроциты, построив график скорости диффузия (микромоль в час) в зависимости от внешней концентрации глюкозы (ммоль/см 3 ). Для облегчения диффузия, V max = 500 микромолей в час и K = 1,5 ммоль/см 3 . За простая диффузия, А х Р 3 см 3 /час.
Источники: Дарнелл, Дж. , Х. Лодиш и Д. Балтимор. 1986. Молекулярно-клеточная биология. Scientific American Books, Inc., Нью-Йорк
, Х. Лодиш и Д. Балтимор. 1986. Молекулярно-клеточная биология. Scientific American Books, Inc., Нью-Йорк
LeFevre, PG 1961. Транспорт сахара в красной крови. клетка: отношения структура-активность в субстратах и антагонисты. Фармакологический обзор 13 :39-70.
Copyright 1999 M. Beals, L. Gross, S. Harrell
Транспорт малых молекул – клетка
Внутренний состав клетки сохраняется, поскольку плазматическая мембрана избирательно проницаема для малых молекул. Большинство биологических молекул не способны диффундировать через фосфолипидный бислой, поэтому плазматическая мембрана образует барьер, блокирующий свободный обмен молекул между цитоплазмой и внешней средой клетки. Специфические транспортные белки (белки-носители и белковые каналы) опосредуют избирательное прохождение малых молекул через мембрану, позволяя клетке контролировать состав своей цитоплазмы.
Пассивная диффузия
Простейший механизм пересечения молекулами плазматической мембраны — пассивная диффузия. Во время пассивной диффузии молекула просто растворяется в бислое фосфолипидов, диффундирует через него, а затем растворяется в водном растворе на другой стороне мембраны. Никакие мембранные белки не участвуют, и направление транспорта определяется просто относительными концентрациями молекулы внутри и снаружи клетки. Чистый поток молекул всегда направлен вниз по градиенту их концентрации — из отсека с высокой концентрацией в отсек с более низкой концентрацией молекулы.
Во время пассивной диффузии молекула просто растворяется в бислое фосфолипидов, диффундирует через него, а затем растворяется в водном растворе на другой стороне мембраны. Никакие мембранные белки не участвуют, и направление транспорта определяется просто относительными концентрациями молекулы внутри и снаружи клетки. Чистый поток молекул всегда направлен вниз по градиенту их концентрации — из отсека с высокой концентрацией в отсек с более низкой концентрацией молекулы.
Таким образом, пассивная диффузия представляет собой неселективный процесс, при котором любая молекула, способная растворяться в двойном слое фосфолипидов, способна пересекать плазматическую мембрану и уравновешиваться между внутренней и внешней частью клетки. Важно отметить, что только небольшие, относительно гидрофобные молекулы способны диффундировать через бислой фосфолипидов со значительной скоростью. Так, газы (такие как O 2 и CO 2 ), гидрофобные молекулы (такие как бензол) и небольшие полярные, но незаряженные молекулы (такие как H 2 O и этанол) способны диффундировать через плазматическую мембрану. Однако другие биологические молекулы не способны растворяться в гидрофобной внутренней части двойного слоя фосфолипидов. Следовательно, более крупные незаряженные полярные молекулы, такие как глюкоза, не могут пересечь плазматическую мембрану путем пассивной диффузии, как и заряженные молекулы любого размера (включая небольшие ионы, такие как H + , Na + , K + и Cl). – ). Прохождение этих молекул через мембрану вместо этого требует активности специфических транспортных и канальных белков, которые, таким образом, контролируют движение большинства биологических молекул в клетку и из нее.
Однако другие биологические молекулы не способны растворяться в гидрофобной внутренней части двойного слоя фосфолипидов. Следовательно, более крупные незаряженные полярные молекулы, такие как глюкоза, не могут пересечь плазматическую мембрану путем пассивной диффузии, как и заряженные молекулы любого размера (включая небольшие ионы, такие как H + , Na + , K + и Cl). – ). Прохождение этих молекул через мембрану вместо этого требует активности специфических транспортных и канальных белков, которые, таким образом, контролируют движение большинства биологических молекул в клетку и из нее.
Рисунок 12.15
Проницаемость фосфолипидных бислоев. Газы, гидрофобные молекулы и небольшие полярные незаряженные молекулы могут диффундировать через бислои фосфолипидов. Более крупные полярные молекулы и заряженные молекулы не могут.
Облегченная диффузия и белки-носители
Облегченная диффузия, как и пассивная, включает движение молекул в направлении, определяемом их относительными концентрациями внутри и снаружи клетки. Никакого внешнего источника энергии не предусмотрено, поэтому молекулы движутся через мембрану в направлении, определяемом их градиентами концентрации и, в случае заряженных молекул, электрическим потенциалом на мембране. Однако облегченная диффузия отличается от пассивной диффузии тем, что транспортируемые молекулы не растворяются в двойном слое фосфолипидов. Вместо этого их прохождение опосредуется белками, которые позволяют транспортируемым молекулам пересекать мембрану, не взаимодействуя непосредственно с ее гидрофобной внутренней частью. Таким образом, облегченная диффузия позволяет полярным и заряженным молекулам, таким как углеводы, аминокислоты, нуклеозиды и ионы, пересекать плазматическую мембрану.
Никакого внешнего источника энергии не предусмотрено, поэтому молекулы движутся через мембрану в направлении, определяемом их градиентами концентрации и, в случае заряженных молекул, электрическим потенциалом на мембране. Однако облегченная диффузия отличается от пассивной диффузии тем, что транспортируемые молекулы не растворяются в двойном слое фосфолипидов. Вместо этого их прохождение опосредуется белками, которые позволяют транспортируемым молекулам пересекать мембрану, не взаимодействуя непосредственно с ее гидрофобной внутренней частью. Таким образом, облегченная диффузия позволяет полярным и заряженным молекулам, таким как углеводы, аминокислоты, нуклеозиды и ионы, пересекать плазматическую мембрану.
Обычно различают два класса белков, которые опосредуют облегченную диффузию: белки-носители и белки-каналы. Белки-переносчики связывают определенные молекулы для транспортировки на одной стороне мембраны. Затем они претерпевают конформационные изменения, которые позволяют молекуле пройти через мембрану и выйти на другую сторону. Напротив, канальные белки (см. следующий раздел) образуют открытые поры через мембрану, обеспечивая свободную диффузию любой молекулы соответствующего размера и заряда.
Напротив, канальные белки (см. следующий раздел) образуют открытые поры через мембрану, обеспечивая свободную диффузию любой молекулы соответствующего размера и заряда.
Белки-носители отвечают за облегченную диффузию сахаров, аминокислот и нуклеозидов через плазматические мембраны большинства клеток. Поглощение глюкозы, которая служит основным источником метаболической энергии, является одной из наиболее важных транспортных функций плазматической мембраны, а транспортер глюкозы является хорошо изученным примером белка-переносчика. Транспортер глюкозы первоначально был идентифицирован как белок массой 55 кД в эритроцитах человека, в которых он составляет примерно 5% от общего белка мембраны. Последующее выделение и анализ последовательности клона кДНК выявили, что переносчик глюкозы имеет 12 α-спиральных трансмембранных сегментов — структура, типичная для многих белков-переносчиков (12). Эти трансмембранные α-спирали содержат преимущественно гидрофобные аминокислоты, но некоторые из них также содержат полярные аминокислотные остатки, которые, как считается, образуют сайт связывания глюкозы внутри белка.
Рисунок 12.16
Структура переносчика глюкозы. Транспортер глюкозы имеет 12 трансмембранных α-спиралей. Полярные аминокислотные остатки, расположенные внутри фосфолипидного бислоя, обозначены темно-фиолетовыми кружками. (Адаптировано из G. I. Bell, C. F. Burant, J. Takeda (подробнее…)
Как и для многих мембранных белков, трехмерная структура транспортера глюкозы неизвестна, поэтому молекулярный механизм транспорта глюкозы остается открытым вопросом. Однако кинетические исследования показывают, что переносчик глюкозы функционирует путем чередования двух конформационных состояний (1). В первой конформации сайт связывания глюкозы обращен к внешней стороне клетки. Связывание глюкозы с этим внешним сайтом вызывает конформационные изменения в транспортер, так что сайт связывания глюкозы теперь обращен внутрь клетки.Затем глюкоза может высвобождаться в цитозоль, после чего транспортер возвращается к своей исходной конформации.
Рисунок 12.17
Модель облегченной диффузии глюкозы. Транспортер глюкозы чередуется между двумя конформациями, в которых сайт связывания глюкозы попеременно экспонируется снаружи и внутри клетки. В первой показанной конформации (A) глюкоза (подробнее…)
Транспортер глюкозы чередуется между двумя конформациями, в которых сайт связывания глюкозы попеременно экспонируется снаружи и внутри клетки. В первой показанной конформации (A) глюкоза (подробнее…)
Большинство клеток, включая эритроциты, подвергаются воздействию внеклеточных концентраций глюкозы, которые выше, чем концентрации внутри клетки, поэтому облегченная диффузия приводит к чистому транспорту глюкозы внутрь . Как только глюкоза поглощается этими клетками, она быстро метаболизируется, поэтому внутриклеточные концентрации глюкозы остаются низкими, и глюкоза продолжает транспортироваться в клетку из внеклеточной жидкости. Однако, поскольку конформационные изменения переносчика глюкозы обратимы, глюкоза может транспортироваться в противоположном направлении, просто обращая шаги в . Такой обратный поток имеет место, например, в клетках печени, в которых синтезируется глюкоза и высвобождается в кровоток.
Ионные каналы
В отличие от белков-носителей, канальные белки просто образуют открытые поры в мембране, позволяя небольшим молекулам соответствующего размера и заряда свободно проходить через липидный бислой. Одна группа белков каналов, обсуждавшаяся ранее, — это порины, которые обеспечивают свободное прохождение ионов и небольших полярных молекул через наружные мембраны бактерий (см. ). Канальные белки также обеспечивают прохождение молекул между клетками, соединенными щелевыми контактами, которые обсуждаются далее в этой главе. Плазматические мембраны многих клеток также содержат белки водных каналов (аквапорины), через которые молекулы воды способны пересекать мембрану гораздо быстрее, чем они могут диффундировать через бислой фосфолипидов. Однако наиболее хорошо охарактеризованными белками каналов являются ионные каналы, которые опосредуют прохождение ионов через плазматические мембраны. Хотя ионные каналы присутствуют в мембранах всех клеток, они особенно хорошо изучены в нервах и мышцах, где их регулируемое открытие и закрытие отвечает за передачу электрических сигналов.
Одна группа белков каналов, обсуждавшаяся ранее, — это порины, которые обеспечивают свободное прохождение ионов и небольших полярных молекул через наружные мембраны бактерий (см. ). Канальные белки также обеспечивают прохождение молекул между клетками, соединенными щелевыми контактами, которые обсуждаются далее в этой главе. Плазматические мембраны многих клеток также содержат белки водных каналов (аквапорины), через которые молекулы воды способны пересекать мембрану гораздо быстрее, чем они могут диффундировать через бислой фосфолипидов. Однако наиболее хорошо охарактеризованными белками каналов являются ионные каналы, которые опосредуют прохождение ионов через плазматические мембраны. Хотя ионные каналы присутствуют в мембранах всех клеток, они особенно хорошо изучены в нервах и мышцах, где их регулируемое открытие и закрытие отвечает за передачу электрических сигналов.
Три свойства ионных каналов лежат в основе их функции (). Во-первых, транспортировка по каналам чрезвычайно быстра. Более миллиона ионов в секунду проходят через открытые каналы — скорость потока примерно в тысячу раз превышает скорость транспорта белков-переносчиков. Во-вторых, ионные каналы обладают высокой селективностью, поскольку узкие поры в канале ограничивают прохождение ионов соответствующего размера и заряда. Таким образом, специфические белки каналов обеспечивают прохождение Na + , K + , Ca 2+ и Cl – через мембрану. В-третьих, большинство ионных каналов открыты не постоянно. Вместо этого открытие ионных каналов регулируется «воротами», которые кратковременно открываются в ответ на определенные раздражители. Некоторые каналы (называемые лиганд-зависимыми каналами ) открываются в ответ на связывание нейротрансмиттеров или других сигнальных молекул; другие ( потенциалзависимых канала ) открываются в ответ на изменения электрического потенциала на плазматической мембране.
Более миллиона ионов в секунду проходят через открытые каналы — скорость потока примерно в тысячу раз превышает скорость транспорта белков-переносчиков. Во-вторых, ионные каналы обладают высокой селективностью, поскольку узкие поры в канале ограничивают прохождение ионов соответствующего размера и заряда. Таким образом, специфические белки каналов обеспечивают прохождение Na + , K + , Ca 2+ и Cl – через мембрану. В-третьих, большинство ионных каналов открыты не постоянно. Вместо этого открытие ионных каналов регулируется «воротами», которые кратковременно открываются в ответ на определенные раздражители. Некоторые каналы (называемые лиганд-зависимыми каналами ) открываются в ответ на связывание нейротрансмиттеров или других сигнальных молекул; другие ( потенциалзависимых канала ) открываются в ответ на изменения электрического потенциала на плазматической мембране.
Рисунок 12.18
Модель ионного канала. В закрытой конформации поток ионов блокируется воротами. Открытие ворот позволяет ионам быстро проходить через канал. Канал содержит узкую пору, которая ограничивает прохождение ионов соответствующего размера (подробнее…)
В закрытой конформации поток ионов блокируется воротами. Открытие ворот позволяет ионам быстро проходить через канал. Канал содержит узкую пору, которая ограничивает прохождение ионов соответствующего размера (подробнее…)
Фундаментальная роль ионных каналов в передаче электрических импульсов была выяснена в серии элегантных экспериментов, описанных Аланом Ходжкиным и Эндрю Хаксли. в 1952 году. Эти исследователи использовали в качестве модели гигантские нервные клетки кальмара. Аксоны этих гигантских нейронов имеют диаметр около 1 мм, что позволяет вводить электроды и измерять изменения мембранного потенциала, происходящие при передаче нервных импульсов. Используя этот подход, Ходжкин и Хаксли продемонстрировали, что эти изменения мембранного потенциала являются результатом регулируемого открытия и закрытия Na + и К + каналы в плазматической мембране. Впоследствии стало возможным изучать активность отдельных ионных каналов с помощью методики patch-clamp , разработанной Erwin Neher и Bert Sakmann в 1976 г. (). В этом методе микропипетка с диаметром наконечника около 1 мкм используется для выделения небольшого участка мембраны, что позволяет анализировать поток ионов через один канал и значительно повышает точность, с которой можно определить активность ионных каналов. учился.
(). В этом методе микропипетка с диаметром наконечника около 1 мкм используется для выделения небольшого участка мембраны, что позволяет анализировать поток ионов через один канал и значительно повышает точность, с которой можно определить активность ионных каналов. учился.
Рисунок 12.19
Метод накладного зажима. На кончике микропипетки выделяют небольшой участок мембраны. Затем внутри пипетки можно применять стимулы, что позволяет измерить поведение захваченного канала. (Адаптировано из E. Neher and B. Sakmann, 1992. (подробнее…)
Поток ионов через мембранные каналы зависит от установления ионных градиентов через плазматическую мембрану. Все клетки, включая нервные и мышечные, содержат ионных насосов (обсуждается в следующем разделе), которые используют энергию, полученную в результате гидролиза АТФ, для активного транспорта ионов через плазматическую мембрану.В результате ионный состав цитоплазмы существенно отличается от внеклеточной жидкости ().Например, Na + активно выкачивается из клеток, а К + закачивается. Таким образом, в аксоне кальмара концентрация Na + примерно в 10 раз выше во внеклеточной жидкости, чем внутри клетки, тогда как концентрация К + примерно в 20 раз выше в цитозоле, чем в окружающей среде.
Таким образом, в аксоне кальмара концентрация Na + примерно в 10 раз выше во внеклеточной жидкости, чем внутри клетки, тогда как концентрация К + примерно в 20 раз выше в цитозоле, чем в окружающей среде.
Таблица 12.1
Концентрации внеклеточных и внутриклеточных ионов.
Поскольку ионы электрически заряжены, их перенос приводит к возникновению электрического градиента на плазматической мембране. В покоящихся аксонах кальмара существует электрический потенциал около 60 мВ через плазматическую мембрану, при этом внутренняя часть клетки отрицательная по отношению к внешней (). Этот электрический потенциал возникает как от ионных насосов, так и от потока ионов через каналы, открытые в плазматической мембране покоящейся клетки. Плазматическая мембрана аксонов покоящегося кальмара содержит открытые K + , поэтому он более проницаем для K + , чем для Na + или других ионов. Следовательно, поток К + вносит наибольший вклад в мембранный потенциал покоя.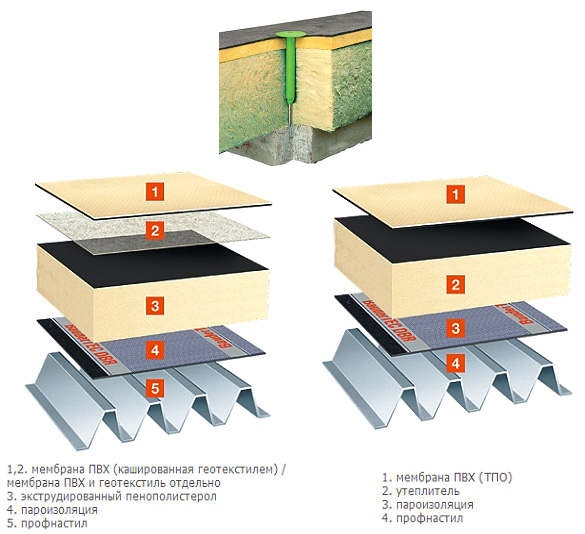
Рис. 12.20
Ионные градиенты и мембранный потенциал покоя аксона гигантского кальмара. Показаны только концентрации Na + и K + , поскольку именно эти ионы участвуют в передаче нервных импульсов. Na + выкачивается из клетки, а K + закачивается, (подробнее…)
Как обсуждалось в главе 10, поток ионов через мембрану управляется как концентрацией, так и компонентами напряжения электрохимического градиента. Например, концентрация K + внутри аксона кальмара в 20 раз выше, чем во внеклеточной жидкости, что приводит к выходу K + из клетки. Однако, поскольку K + заряжен положительно, этот выход K + из клетки генерирует электрический потенциал через мембрану, при этом внутренняя часть клетки становится отрицательно заряженной. Этот мембранный потенциал противостоит продолжающемуся потоку K + из клетки, и система приближается к равновесному состоянию, при котором мембранный потенциал уравновешивает градиент концентрации К + .
Количественно связь между концентрацией ионов и мембранным потенциалом определяется уравнением Нернста:
где В – равновесный потенциал в вольтах, R – газовая постоянная, T – абсолютная температура, z – заряд иона, F — постоянная Фарадея, а C o и C i — концентрации иона снаружи и внутри клетки соответственно. Равновесный потенциал существует отдельно для каждого иона, а мембранный потенциал определяется потоком всех ионов, пересекающих плазматическую мембрану. Однако, поскольку покоящиеся аксоны кальмаров более проницаемы для K + , чем для Na + или других ионов (включая Cl – ), мембранный потенциал покоя (-60 мВ) близок к равновесному потенциалу, определяемому внутриклеточным и внеклеточный K + концентрации (-75 мВ).
Когда нервные импульсы ( потенциалы действия ) проходят по аксонам, мембрана деполяризуется (). Мембранный потенциал изменяется от -60 мВ до примерно +30 мВ менее чем за миллисекунду, после чего снова становится отрицательным и возвращается к своему значению покоя. Эти изменения являются результатом быстрого последовательного открытия и закрытия потенциалзависимых каналов Na + и K + . Относительно небольшие начальные изменения мембранного потенциала (от -60 до примерно -40 мВ) приводят к быстрому открытию Na + канала. Это позволяет Na + поступать в клетку под действием как градиента его концентрации, так и мембранного потенциала. Внезапное поступление Na + приводит к сильному изменению мембранного потенциала, который увеличивается почти до +30 мВ, приближаясь к равновесному потенциалу Na + примерно +50 мВ. В это время каналы Na + инактивируются, а потенциалзависимые каналы K + открываются, существенно повышая проницаемость мембраны для K + . Затем K + быстро вытекает из клетки под действием как мембранного потенциала, так и градиента концентрации K + , что приводит к быстрому снижению мембранного потенциала примерно до -75 мВ (равновесный потенциал K + ).
Эти изменения являются результатом быстрого последовательного открытия и закрытия потенциалзависимых каналов Na + и K + . Относительно небольшие начальные изменения мембранного потенциала (от -60 до примерно -40 мВ) приводят к быстрому открытию Na + канала. Это позволяет Na + поступать в клетку под действием как градиента его концентрации, так и мембранного потенциала. Внезапное поступление Na + приводит к сильному изменению мембранного потенциала, который увеличивается почти до +30 мВ, приближаясь к равновесному потенциалу Na + примерно +50 мВ. В это время каналы Na + инактивируются, а потенциалзависимые каналы K + открываются, существенно повышая проницаемость мембраны для K + . Затем K + быстро вытекает из клетки под действием как мембранного потенциала, так и градиента концентрации K + , что приводит к быстрому снижению мембранного потенциала примерно до -75 мВ (равновесный потенциал K + ). Затем потенциалзависимые каналы K + инактивируются, и мембранный потенциал возвращается к уровню покоя -60 мВ, определяемому потоком K + и других ионов через каналы, которые остаются открытыми в нестимулированных клетках.
Затем потенциалзависимые каналы K + инактивируются, и мембранный потенциал возвращается к уровню покоя -60 мВ, определяемому потоком K + и других ионов через каналы, которые остаются открытыми в нестимулированных клетках.
Рис. 12.21
Мембранный потенциал и ионные каналы во время потенциала действия. (A) Изменения мембранного потенциала в одной точке гигантского аксона кальмара после стимула. E Na и E K – равновесные потенциалы Na + и K + соответственно. (B) Мембранный потенциал (подробнее…)
Деполяризация соседних областей плазматической мембраны позволяет потенциалам действия проходить по длине аксонов нервных клеток в виде электрических сигналов, что приводит к быстрой передаче нервных импульсов на большие расстояния. Например, аксоны двигательных нейронов человека могут иметь длину более метра. Прибытие потенциалов действия на окончание большинства нейронов сигнализирует о высвобождении нейротрансмиттеров, таких как ацетилхолин, которые передают сигналы между клетками в синапсах. Нейротрансмиттеры, высвобождаемые из пресинаптических клеток, связываются с рецепторами на мембранах постсинаптических клеток, где они действуют, открывая управляемые лигандом ионные каналы. Одним из наиболее хорошо охарактеризованных таких каналов является ацетилхолиновый рецептор мышечных клеток. Связывание ацетилхолина открывает канал, проницаемый как для Na + и К +. Это обеспечивает быстрый приток Na + , который деполяризует мембрану мышечной клетки и запускает потенциал действия. Затем потенциал действия приводит к открытию потенциалзависимых каналов Ca 2+ , что приводит к увеличению внутриклеточного количества Ca 2+ , что сигнализирует о сокращении (see ).
Нейротрансмиттеры, высвобождаемые из пресинаптических клеток, связываются с рецепторами на мембранах постсинаптических клеток, где они действуют, открывая управляемые лигандом ионные каналы. Одним из наиболее хорошо охарактеризованных таких каналов является ацетилхолиновый рецептор мышечных клеток. Связывание ацетилхолина открывает канал, проницаемый как для Na + и К +. Это обеспечивает быстрый приток Na + , который деполяризует мембрану мышечной клетки и запускает потенциал действия. Затем потенциал действия приводит к открытию потенциалзависимых каналов Ca 2+ , что приводит к увеличению внутриклеточного количества Ca 2+ , что сигнализирует о сокращении (see ).
Рисунок 12.22
Передача сигналов за счет высвобождения нейротрансмиттера в синапсе. Прибытие нервного импульса на окончание нейрона сигнализирует о слиянии синаптических пузырьков с плазматической мембраной, что приводит к высвобождению нейротрансмиттера из пресинаптической клетки в (далее. ..)
..)
Ацетилхолиновый рецептор, первоначально выделенный из электрического органа скатов Torpedo в 1970-х годах, является прототипом лиганд-управляемых каналов. Рецептор состоит из пяти субъединиц, расположенных в виде цилиндра в мембране (). Считается, что в закрытом состоянии поры канала блокируются боковыми цепями гидрофобных аминокислот. Связывание ацетилхолина вызывает конформационные изменения в рецепторе, так что эти гидрофобные боковые цепи смещаются из канала, открывая пору, через которую проходят положительно заряженные ионы, включая Na + и К +. Однако канал остается непроницаемым для отрицательно заряженных ионов, таких как Cl –, поскольку он выстлан отрицательно заряженными аминокислотами.
Рисунок 12.23
Модель рецептора ацетилхолина. Рецептор состоит из пяти субъединиц, расположенных вокруг центральной поры. Связывание ацетилхолина с участком во внеклеточной области рецептора вызывает аллостерические изменения, которые открывают ворота канала.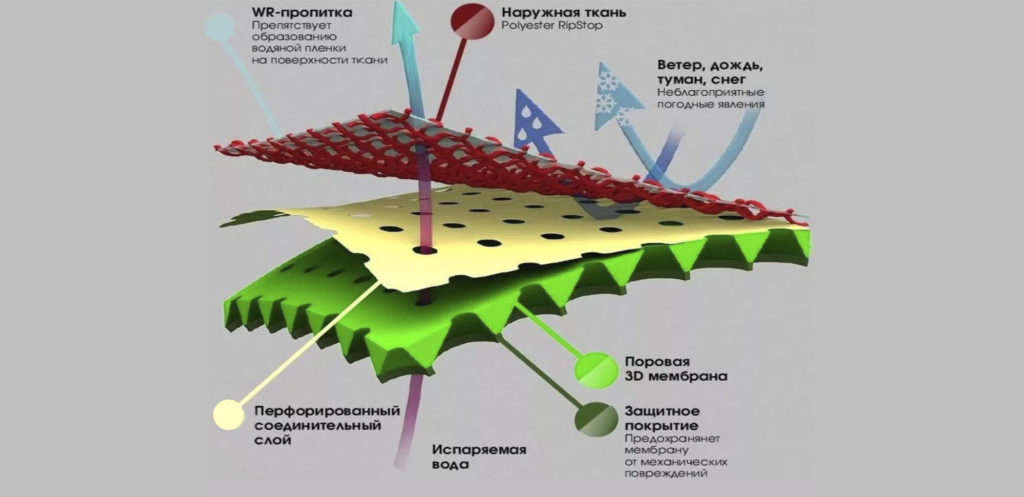 Канал (подробнее…)
Канал (подробнее…)
Большую степень ионной селективности проявляют потенциалзависимые каналы Na + и K + . Каналы Na + более чем в десять раз более проницаемы для Na + , чем для K + , тогда как каналы K + более чем в тысячу раз более проницаемы для K + , чем для Na + . Селективность канала Na + может быть объяснена, по крайней мере, частично, на основе узких пор, которые действуют как размерный фильтр. Ионный радиус Na + (0,95 Å) меньше, чем у K + (1,33 Å), и считается, что пора канала Na + достаточно узкая, чтобы препятствовать прохождению ионов K + или более крупных ( ).
Рисунок 12.24
Ионная селективность каналов Na + . Узкая пора позволяет проходить Na + , связанным с одной молекулой воды, но препятствует прохождению ионов K + или более крупных ионов.
K + каналы также имеют узкие поры, препятствующие прохождению более крупных ионов. Однако, поскольку Na + имеет меньший ионный радиус, это не объясняет избирательной проницаемости этих каналов для K + . Селективность канала K + основана на другом механизме, который был выяснен при определении трехмерной структуры канала K + с помощью рентгеновской кристаллографии в 1998 г. (). Пора канала содержит фильтр с узкой селективностью, выстланный атомами карбонильного кислорода (C=O) из основной цепи полипептида. Когда К 9Ион 0061 + поступает в фильтр селективности, взаимодействия с этими карбонильными атомами кислорода вытесняют молекулы воды, с которыми связан K + , позволяя обезвоженному K + проходить через поры. Напротив, обезвоженный Na + слишком мал, чтобы взаимодействовать с этими карбонильными атомами кислорода в селективном фильтре, который удерживается жестко открытым. Следовательно, Na + остается связанным с молекулами воды в гидратированном комплексе, который слишком велик, чтобы пройти через канал.
Однако, поскольку Na + имеет меньший ионный радиус, это не объясняет избирательной проницаемости этих каналов для K + . Селективность канала K + основана на другом механизме, который был выяснен при определении трехмерной структуры канала K + с помощью рентгеновской кристаллографии в 1998 г. (). Пора канала содержит фильтр с узкой селективностью, выстланный атомами карбонильного кислорода (C=O) из основной цепи полипептида. Когда К 9Ион 0061 + поступает в фильтр селективности, взаимодействия с этими карбонильными атомами кислорода вытесняют молекулы воды, с которыми связан K + , позволяя обезвоженному K + проходить через поры. Напротив, обезвоженный Na + слишком мал, чтобы взаимодействовать с этими карбонильными атомами кислорода в селективном фильтре, который удерживается жестко открытым. Следовательно, Na + остается связанным с молекулами воды в гидратированном комплексе, который слишком велик, чтобы пройти через канал.
Рисунок 12.25
Избирательность каналов K + . Канал К + содержит фильтр узкой селективности, выстланный карбонильными атомами кислорода (С=О). Поры достаточно широки, чтобы обеспечить прохождение обезвоженного K + , из которого все связанные молекулы воды были вытеснены в виде (далее…)
Потенциалзависимых Na + , K + и Ca 2+ канала принадлежат к большому семейству родственных белков (2). Например, последовательность генома C. elegans выявил почти 200 генов, кодирующих ионные каналы, которые, по-видимому, необходимы, чтобы играть различные роли в клеточной передаче сигналов. Каналы K + состоят из четырех идентичных субъединиц, каждая из которых содержит две или шесть трансмембранных α-спиралей. Каналы Na + и Ca 2+ состоят из одной полипептидной цепи, но каждый полипептид содержит четыре повторяющихся домена, соответствующих субъединицам каналов K + . Запирание напряжения опосредовано одной из трансмембранных α-спиралей, которая содержит несколько положительно заряженных аминокислот. Мембранная деполяризация индуцирует перемещение этих положительных зарядов наружу клетки, сдвигая положение этого трансмембранного сегмента и открывая канал. Быстрая инактивация Na + и К + каналы при распространении потенциалов действия затем опосредованы цитоплазматическими участками полипептидной цепи, которые связываются с цитоплазматическим устьем поры канала и препятствуют дальнейшему току ионов ().
Запирание напряжения опосредовано одной из трансмембранных α-спиралей, которая содержит несколько положительно заряженных аминокислот. Мембранная деполяризация индуцирует перемещение этих положительных зарядов наружу клетки, сдвигая положение этого трансмембранного сегмента и открывая канал. Быстрая инактивация Na + и К + каналы при распространении потенциалов действия затем опосредованы цитоплазматическими участками полипептидной цепи, которые связываются с цитоплазматическим устьем поры канала и препятствуют дальнейшему току ионов ().
Рисунок 12.26
Структуры потенциалзависимых катионных каналов. Каналы K + , Na + и Ca 2+ принадлежат к семейству родственных белков. Канал К + образован объединением четырех идентичных субъединиц, одна из которых показана. На 9Канал 0061 + состоит из одного полипептида (подробнее…)
Рисунок 12.27
Инактивация каналов K + и Na + . После потенциалзависимого открытия каналы K + и Na + быстро инактивируются за счет связывания цитоплазматических частей полипептидных цепей с порой. Для канала K + инактивация опосредована шаровой цепью (подробнее…)
После потенциалзависимого открытия каналы K + и Na + быстро инактивируются за счет связывания цитоплазматических частей полипептидных цепей с порой. Для канала K + инактивация опосредована шаровой цепью (подробнее…)
Широкий спектр ионных каналов (включая Ca 2+ и Cl – каналы) реагируют на разные нейротрансмиттеры или открываются и закрываются с разной кинетикой после деполяризации мембраны. Согласованные действия этих многочисленных каналов ответственны за сложность передачи сигналов в нервной системе. Более того, как будет показано в следующей главе, роль ионных каналов не ограничивается электрически возбудимыми нервными и мышечными клетками; они также играют важную роль в передаче сигналов в других типах клеток. Таким образом, регулируемое открытие и закрытие ионных каналов обеспечивает клетки чувствительным и универсальным механизмом реагирования на различные стимулы окружающей среды.
Активный транспорт, управляемый гидролизом АТФ
Чистый поток молекул за счет облегченной диффузии либо через белки-носители, либо через белки-каналы всегда энергетически направлен вниз в направлении, определяемом электрохимическими градиентами через мембрану. Однако во многих случаях клетка должна транспортировать молекулы против градиента их концентрации. При активном транспорте энергия, обеспечиваемая другой связанной реакцией (например, гидролизом АТФ), используется для перемещения молекул в гору в энергетически невыгодном направлении.
Однако во многих случаях клетка должна транспортировать молекулы против градиента их концентрации. При активном транспорте энергия, обеспечиваемая другой связанной реакцией (например, гидролизом АТФ), используется для перемещения молекул в гору в энергетически невыгодном направлении.
Ионные насосы, отвечающие за поддержание градиента ионов через плазматическую мембрану, являются важными примерами активного транспорта, управляемого непосредственно гидролизом АТФ. Как обсуждалось ранее (см.), концентрация Na + примерно в десять раз выше снаружи, чем внутри клеток, тогда как концентрация K + выше внутри, чем снаружи. Эти градиенты ионов поддерживаются насосом Na + -K + (также называемая Na + -K + АТФаза ), которая использует энергию, полученную в результате гидролиза АТФ, для транспорта Na + и K + против их электрохимических градиентов. Этот процесс является результатом конформационных изменений в помпе, вызванных АТФ. Во-первых, ионы Na + связываются с центрами высокого сродства внутри клетки. Это связывание стимулирует гидролиз АТФ и фосфорилирование помпы, вызывая конформационные изменения, которые обнажают Na + – связывает сайты снаружи клетки и снижает их сродство к Na + . Следовательно, связанный Na + высвобождается во внеклеточную жидкость. В то же время на клеточной поверхности экспонируются высокоаффинные сайты связывания К + -. Связывание внеклеточного K + с этими сайтами затем стимулирует гидролиз фосфатной группы, связанной с насосом, что вызывает второе конформационное изменение, открывая сайты связывания K + цитозолю и снижая их аффинность связывания, так что K + высвобождается внутри клетки. Насос имеет три сайта связывания для Na + и два для K + , поэтому каждый цикл переносит через плазматическую мембрану три Na + и два K + за счет одной молекулы АТФ.
Этот процесс является результатом конформационных изменений в помпе, вызванных АТФ. Во-первых, ионы Na + связываются с центрами высокого сродства внутри клетки. Это связывание стимулирует гидролиз АТФ и фосфорилирование помпы, вызывая конформационные изменения, которые обнажают Na + – связывает сайты снаружи клетки и снижает их сродство к Na + . Следовательно, связанный Na + высвобождается во внеклеточную жидкость. В то же время на клеточной поверхности экспонируются высокоаффинные сайты связывания К + -. Связывание внеклеточного K + с этими сайтами затем стимулирует гидролиз фосфатной группы, связанной с насосом, что вызывает второе конформационное изменение, открывая сайты связывания K + цитозолю и снижая их аффинность связывания, так что K + высвобождается внутри клетки. Насос имеет три сайта связывания для Na + и два для K + , поэтому каждый цикл переносит через плазматическую мембрану три Na + и два K + за счет одной молекулы АТФ.
Рисунок 12.28
Модель для работы насоса Na + -K + .
Важность помпы Na + -K + подтверждается тем фактом, что она потребляет почти 25% АТФ, используемого многими животными клетками. Одна из важнейших ролей Na + и К + градиенты, установленные насосом, — это распространение электрических сигналов в нерве и мышце. Как будет сказано ниже, градиент Na + , создаваемый помпой, также используется для обеспечения активного транспорта множества других молекул. Еще одна важная роль насоса Na + -K + в большинстве клеток животных заключается в поддержании осмотического баланса и объема клетки. Цитоплазма содержит высокую концентрацию органических молекул, включая макромолекулы, аминокислоты, сахара и нуклеотиды. В отсутствие противовеса это будет стимулировать поток воды внутрь за счет осмоса, что, если его не остановить, приведет к набуханию и, в конечном итоге, к разрыву клетки. Необходимый противовес обеспечивается градиентами ионов, установленными Na + -К + насос (). В частности, насос устанавливает более высокую концентрацию Na + снаружи, чем внутри клетки. Как уже обсуждалось, поток K + через открытые каналы дополнительно устанавливает электрический потенциал через плазматическую мембрану. Этот мембранный потенциал, в свою очередь, выталкивает Cl – из клетки, поэтому концентрация Cl – (как и Na + ) примерно в десять раз выше во внеклеточной жидкости, чем в цитоплазме. Эти различия в концентрациях ионов уравновешивают высокие концентрации органических молекул внутри клеток, выравнивая осмотическое давление и предотвращая чистый приток воды.
Необходимый противовес обеспечивается градиентами ионов, установленными Na + -К + насос (). В частности, насос устанавливает более высокую концентрацию Na + снаружи, чем внутри клетки. Как уже обсуждалось, поток K + через открытые каналы дополнительно устанавливает электрический потенциал через плазматическую мембрану. Этот мембранный потенциал, в свою очередь, выталкивает Cl – из клетки, поэтому концентрация Cl – (как и Na + ) примерно в десять раз выше во внеклеточной жидкости, чем в цитоплазме. Эти различия в концентрациях ионов уравновешивают высокие концентрации органических молекул внутри клеток, выравнивая осмотическое давление и предотвращая чистый приток воды.
Рис. 12.29
Градиенты ионов через плазматическую мембрану типичной клетки млекопитающих. Концентрации Na + и Cl – выше снаружи, чем внутри клетки, тогда как концентрация К + выше внутри, чем снаружи. Низкие концентрации Na + и Cl – уравновешивают (подробнее…)
Низкие концентрации Na + и Cl – уравновешивают (подробнее…)
Активный транспорт Ca 2+ через плазматическую мембрану управляется насосом Ca 2+ , который структурно родственен до На + -K + насос и аналогичным образом питается от гидролиза АТФ. Насос Ca 2+ транспортирует Ca 2+ из клетки, поэтому внутриклеточные концентрации Ca 2+ чрезвычайно низки: приблизительно 0,1 мкМ, по сравнению с внеклеточными концентрациями около 1 м M . Эта низкая внутриклеточная концентрация Ca 2+ делает клетку чувствительной к небольшому увеличению внутриклеточного уровня Ca 2+ . Такое временное увеличение внутриклеточного Ca 2+ играют важную роль в клеточной передаче сигналов, как уже отмечалось в отношении мышечного сокращения (см. ) и обсуждается далее в следующей главе.
Аналогичные ионные насосы в плазматических мембранах бактерий, дрожжей и растительных клеток ответственны за активный транспорт H + из клетки. Кроме того, H + активно выкачиваются из клеток, выстилающих желудок, что приводит к повышению кислотности желудочного сока. Структурно различные помпы ответственны за активный транспорт H + на лизосомы и эндосомы (см.). Тем не менее, третий тип насоса H + иллюстрируется АТФ-синтазами митохондрий и хлоропластов: в этих случаях насосы можно рассматривать как работающие в обратном направлении, когда движение ионов по электрохимическому градиенту используется для управления синтезом АТФ.
Кроме того, H + активно выкачиваются из клеток, выстилающих желудок, что приводит к повышению кислотности желудочного сока. Структурно различные помпы ответственны за активный транспорт H + на лизосомы и эндосомы (см.). Тем не менее, третий тип насоса H + иллюстрируется АТФ-синтазами митохондрий и хлоропластов: в этих случаях насосы можно рассматривать как работающие в обратном направлении, когда движение ионов по электрохимическому градиенту используется для управления синтезом АТФ.
Самое большое семейство мембранных транспортеров состоит из транспортеров ABC, названных так потому, что они характеризуются высококонсервативными АТФ-связывающими доменами или A TP- b inding c активы (). Как в прокариотических, так и в эукариотических клетках идентифицировано более сотни членов семейства. У бактерий транспортеры ABC используют энергию, полученную в результате гидролиза АТФ, для транспорта широкого спектра молекул, включая ионы, сахара и аминокислоты. В эукариотических клетках был обнаружен первый переносчик ABC как продукт гена (называемого геном множественной лекарственной устойчивости, или mdr ), который делает раковые клетки устойчивыми к различным лекарствам, используемым в химиотерапии. В настоящее время идентифицированы два переносчика MDR. Обычно они экспрессируются в различных клетках, где они удаляют потенциально токсичные чужеродные соединения. Например, экспрессия переносчика MDR в капиллярных эндотелиальных клетках головного мозга, по-видимому, играет важную роль в защите мозга от токсичных химических веществ. К сожалению, транспортеры MDR часто экспрессируются на высоких уровнях в раковых клетках, где они распознают различные лекарства и выкачивают их из клеток, тем самым делая раковые клетки устойчивыми к широкому спектру химиотерапевтических агентов и создавая серьезное препятствие для успешного лечения рака. .
В эукариотических клетках был обнаружен первый переносчик ABC как продукт гена (называемого геном множественной лекарственной устойчивости, или mdr ), который делает раковые клетки устойчивыми к различным лекарствам, используемым в химиотерапии. В настоящее время идентифицированы два переносчика MDR. Обычно они экспрессируются в различных клетках, где они удаляют потенциально токсичные чужеродные соединения. Например, экспрессия переносчика MDR в капиллярных эндотелиальных клетках головного мозга, по-видимому, играет важную роль в защите мозга от токсичных химических веществ. К сожалению, транспортеры MDR часто экспрессируются на высоких уровнях в раковых клетках, где они распознают различные лекарства и выкачивают их из клеток, тем самым делая раковые клетки устойчивыми к широкому спектру химиотерапевтических агентов и создавая серьезное препятствие для успешного лечения рака. .
Рисунок 12.30
Структура транспортера ABC. Базовая структурная единица транспортеров ABC состоит из шести трансмембранных доменов, за которыми следует АТФ-связывающая кассета. В транспортерах плазматической мембраны (таких как показанный транспортер MDR) две из этих единиц являются (подробнее…)
В транспортерах плазматической мембраны (таких как показанный транспортер MDR) две из этих единиц являются (подробнее…)
Еще одним важным с медицинской точки зрения членом семейства транспортеров ABC является ген, ответственный за муковисцидоз. Хотя он является членом семейства ABC, продукт этого гена (называемый регулятором трансмембранной проводимости при муковисцидозе, или CFTR) функционирует как Cl 9.0061- канал в эпителиальных клетках и дефектный транспорт Cl – характерны для заболевания. Канал CFTR Cl – также необычен тем, что для его открытия, по-видимому, требуется как гидролиз АТФ, так и зависимое от цАМФ фосфорилирование. Структурная основа функции CFTR как регулируемого ионного канала еще предстоит выяснить в ходе будущих исследований.
Активный транспорт, управляемый ионными градиентами
Ионные насосы и АВС-переносчики, рассмотренные в предыдущем разделе, используют энергию, полученную непосредственно в результате гидролиза АТФ, для транспортировки молекул против их электрохимических градиентов. Другие молекулы транспортируются против градиента их концентрации с использованием энергии, полученной не от гидролиза АТФ, а от связанного транспорта второй молекулы в энергетически выгодном направлении. На 9Градиент 0061 + , созданный насосом Na + -K + , обеспечивает источник энергии, который часто используется для обеспечения активного транспорта сахаров, аминокислот и ионов в клетках млекопитающих. Аналогичную роль играют градиенты H + , установленные насосами H + клеток бактерий, дрожжей и растений.
Другие молекулы транспортируются против градиента их концентрации с использованием энергии, полученной не от гидролиза АТФ, а от связанного транспорта второй молекулы в энергетически выгодном направлении. На 9Градиент 0061 + , созданный насосом Na + -K + , обеспечивает источник энергии, который часто используется для обеспечения активного транспорта сахаров, аминокислот и ионов в клетках млекопитающих. Аналогичную роль играют градиенты H + , установленные насосами H + клеток бактерий, дрожжей и растений.
Эпителиальные клетки, выстилающие кишечник, представляют собой хороший пример активного транспорта, управляемого градиентом Na + . Эти клетки используют активные транспортные системы в апикальных областях своих плазматических мембран для захвата пищевых сахаров и аминокислот из просвета кишечника. Поглощение глюкозы, например, осуществляется транспортером, который координированно транспортирует два Na + и одна глюкоза в клетку (). Поток Na + по его электрохимическому градиенту обеспечивает энергию, необходимую для поглощения пищевой глюкозы и накопления высоких концентраций внутриклеточной глюкозы. Затем глюкоза высвобождается в подлежащую соединительную ткань (которая содержит кровеносные капилляры) на базолатеральной поверхности кишечного эпителия, где она транспортируется по градиенту концентрации за счет облегченной диффузии (12). Таким образом, поглощение глюкозы из просвета кишечника и ее высвобождение в кровоток является хорошим примером поляризованной функции эпителиальных клеток, которая является результатом специфической локализации активного транспорта и облегченной диффузии переносчиков в апикальный и базолатеральный домены плазматической мембраны. , соответственно.
Поток Na + по его электрохимическому градиенту обеспечивает энергию, необходимую для поглощения пищевой глюкозы и накопления высоких концентраций внутриклеточной глюкозы. Затем глюкоза высвобождается в подлежащую соединительную ткань (которая содержит кровеносные капилляры) на базолатеральной поверхности кишечного эпителия, где она транспортируется по градиенту концентрации за счет облегченной диффузии (12). Таким образом, поглощение глюкозы из просвета кишечника и ее высвобождение в кровоток является хорошим примером поляризованной функции эпителиальных клеток, которая является результатом специфической локализации активного транспорта и облегченной диффузии переносчиков в апикальный и базолатеральный домены плазматической мембраны. , соответственно.
Рис. 12.31
Активный транспорт глюкозы. Активный транспорт, управляемый градиентом Na + , отвечает за поглощение глюкозы из просвета кишечника. Транспортер координированно связывает и транспортирует в клетку одну глюкозу и две Na + . Транспорт Na + (подробнее…)
Транспорт Na + (подробнее…)
Рис. 12.32
Транспорт глюкозы эпителиальными клетками кишечника. Транспортер в апикальном домене плазматической мембраны отвечает за активное поглощение глюкозы (путем котранспорта с Na + ) из просвета кишечника. В результате пищевая глюкоза поглощается и (подробнее…)
Координированное поглощение глюкозы и Na + является примером симпорта, транспорта двух молекул в одном направлении. Напротив, облегченная диффузия глюкозы является примером унипорта, транспорта только одной молекулы. Активный транспорт также может осуществляться антипортом, при котором две молекулы транспортируются в противоположных направлениях (1). Например, Са 2+ экспортируется из клеток не только насосом Ca 2+ , но и антипортером Na + -Ca 2+ , который транспортирует Na + в клетку и Ca 2+ наружу. Другим примером является обменный белок Na + -H + , который участвует в регуляции внутриклеточного pH. Антипортер Na + -H + связывает транспорт Na + в клетку с экспортом H + , тем самым удаляя избыток H + , образующиеся в результате метаболических реакций и препятствующие закислению цитоплазмы.
Антипортер Na + -H + связывает транспорт Na + в клетку с экспортом H + , тем самым удаляя избыток H + , образующиеся в результате метаболических реакций и препятствующие закислению цитоплазмы.
Рисунок 12.33
Примеры антипорта. Ca 2+ и H + экспортируются из клеток антипортерами, которые связывают их экспорт с энергетически выгодным импортом Na + .
Коробка
Молекулярная медицина: кистозный фиброз.
Транспорт малых молекул — клетка
Внутренний состав клетки сохраняется, поскольку плазматическая мембрана избирательно проницаема для малых молекул. Большинство биологических молекул не способны диффундировать через фосфолипидный бислой, поэтому плазматическая мембрана образует барьер, блокирующий свободный обмен молекул между цитоплазмой и внешней средой клетки. Специфические транспортные белки (белки-носители и белковые каналы) опосредуют избирательное прохождение малых молекул через мембрану, позволяя клетке контролировать состав своей цитоплазмы.
Пассивная диффузия
Простейший механизм пересечения молекулами плазматической мембраны — пассивная диффузия. Во время пассивной диффузии молекула просто растворяется в бислое фосфолипидов, диффундирует через него, а затем растворяется в водном растворе на другой стороне мембраны. Никакие мембранные белки не участвуют, и направление транспорта определяется просто относительными концентрациями молекулы внутри и снаружи клетки. Чистый поток молекул всегда направлен вниз по градиенту их концентрации — из отсека с высокой концентрацией в отсек с более низкой концентрацией молекулы.
Таким образом, пассивная диффузия представляет собой неселективный процесс, при котором любая молекула, способная растворяться в двойном слое фосфолипидов, способна пересекать плазматическую мембрану и уравновешиваться между внутренней и внешней частью клетки. Важно отметить, что только небольшие, относительно гидрофобные молекулы способны диффундировать через бислой фосфолипидов со значительной скоростью.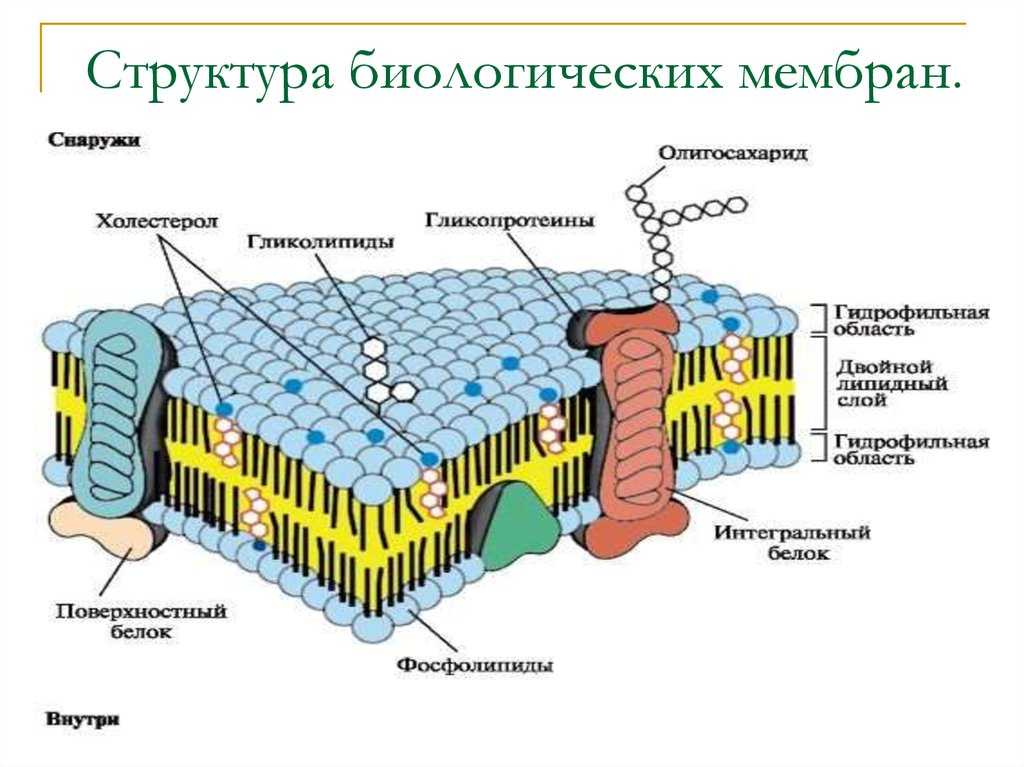 Так, газы (такие как O 2 и CO 2 ), гидрофобные молекулы (такие как бензол) и небольшие полярные, но незаряженные молекулы (такие как H 2 O и этанол) способны диффундировать через плазматическую мембрану. Однако другие биологические молекулы не способны растворяться в гидрофобной внутренней части двойного слоя фосфолипидов. Следовательно, более крупные незаряженные полярные молекулы, такие как глюкоза, не могут пересечь плазматическую мембрану путем пассивной диффузии, как и заряженные молекулы любого размера (включая небольшие ионы, такие как H + , Na + , K + и Cl). – ). Прохождение этих молекул через мембрану вместо этого требует активности специфических транспортных и канальных белков, которые, таким образом, контролируют движение большинства биологических молекул в клетку и из нее.
Так, газы (такие как O 2 и CO 2 ), гидрофобные молекулы (такие как бензол) и небольшие полярные, но незаряженные молекулы (такие как H 2 O и этанол) способны диффундировать через плазматическую мембрану. Однако другие биологические молекулы не способны растворяться в гидрофобной внутренней части двойного слоя фосфолипидов. Следовательно, более крупные незаряженные полярные молекулы, такие как глюкоза, не могут пересечь плазматическую мембрану путем пассивной диффузии, как и заряженные молекулы любого размера (включая небольшие ионы, такие как H + , Na + , K + и Cl). – ). Прохождение этих молекул через мембрану вместо этого требует активности специфических транспортных и канальных белков, которые, таким образом, контролируют движение большинства биологических молекул в клетку и из нее.
Рисунок 12.15
Проницаемость фосфолипидных бислоев. Газы, гидрофобные молекулы и небольшие полярные незаряженные молекулы могут диффундировать через бислои фосфолипидов. Более крупные полярные молекулы и заряженные молекулы не могут.
Более крупные полярные молекулы и заряженные молекулы не могут.
Облегченная диффузия и белки-носители
Облегченная диффузия, как и пассивная, включает движение молекул в направлении, определяемом их относительными концентрациями внутри и снаружи клетки. Никакого внешнего источника энергии не предусмотрено, поэтому молекулы движутся через мембрану в направлении, определяемом их градиентами концентрации и, в случае заряженных молекул, электрическим потенциалом на мембране. Однако облегченная диффузия отличается от пассивной диффузии тем, что транспортируемые молекулы не растворяются в двойном слое фосфолипидов. Вместо этого их прохождение опосредуется белками, которые позволяют транспортируемым молекулам пересекать мембрану, не взаимодействуя непосредственно с ее гидрофобной внутренней частью. Таким образом, облегченная диффузия позволяет полярным и заряженным молекулам, таким как углеводы, аминокислоты, нуклеозиды и ионы, пересекать плазматическую мембрану.
Обычно различают два класса белков, которые опосредуют облегченную диффузию: белки-носители и белки-каналы. Белки-переносчики связывают определенные молекулы для транспортировки на одной стороне мембраны. Затем они претерпевают конформационные изменения, которые позволяют молекуле пройти через мембрану и выйти на другую сторону. Напротив, канальные белки (см. следующий раздел) образуют открытые поры через мембрану, обеспечивая свободную диффузию любой молекулы соответствующего размера и заряда.
Белки-переносчики связывают определенные молекулы для транспортировки на одной стороне мембраны. Затем они претерпевают конформационные изменения, которые позволяют молекуле пройти через мембрану и выйти на другую сторону. Напротив, канальные белки (см. следующий раздел) образуют открытые поры через мембрану, обеспечивая свободную диффузию любой молекулы соответствующего размера и заряда.
Белки-носители отвечают за облегченную диффузию сахаров, аминокислот и нуклеозидов через плазматические мембраны большинства клеток. Поглощение глюкозы, которая служит основным источником метаболической энергии, является одной из наиболее важных транспортных функций плазматической мембраны, а транспортер глюкозы является хорошо изученным примером белка-переносчика. Транспортер глюкозы первоначально был идентифицирован как белок массой 55 кД в эритроцитах человека, в которых он составляет примерно 5% от общего белка мембраны. Последующее выделение и анализ последовательности клона кДНК выявили, что переносчик глюкозы имеет 12 α-спиральных трансмембранных сегментов — структура, типичная для многих белков-переносчиков (12).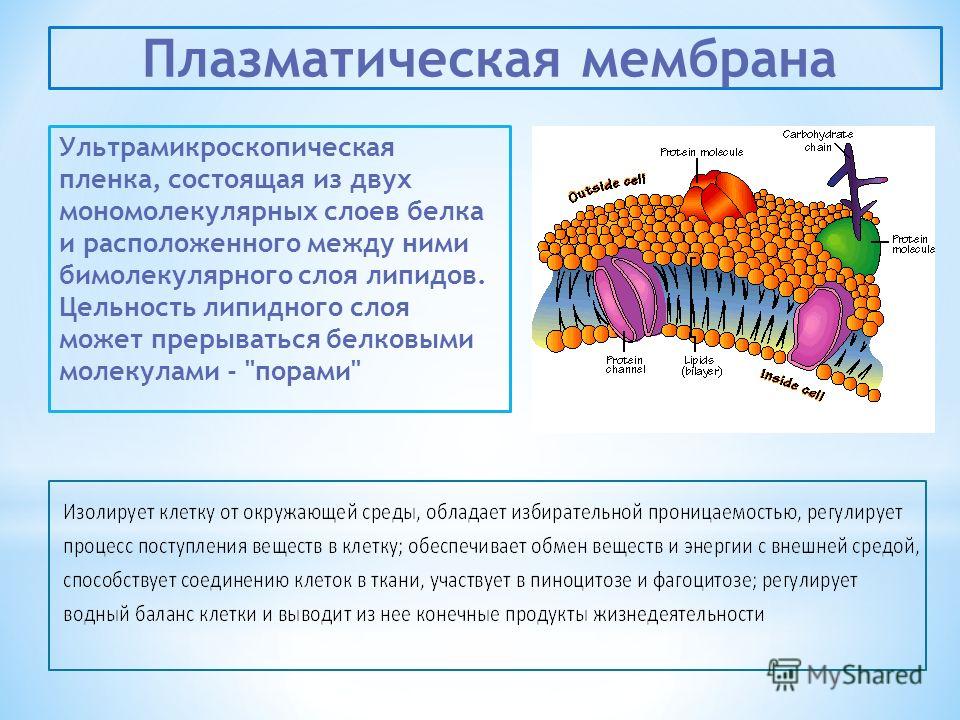 Эти трансмембранные α-спирали содержат преимущественно гидрофобные аминокислоты, но некоторые из них также содержат полярные аминокислотные остатки, которые, как считается, образуют сайт связывания глюкозы внутри белка.
Эти трансмембранные α-спирали содержат преимущественно гидрофобные аминокислоты, но некоторые из них также содержат полярные аминокислотные остатки, которые, как считается, образуют сайт связывания глюкозы внутри белка.
Рисунок 12.16
Структура переносчика глюкозы. Транспортер глюкозы имеет 12 трансмембранных α-спиралей. Полярные аминокислотные остатки, расположенные внутри фосфолипидного бислоя, обозначены темно-фиолетовыми кружками. (Адаптировано из G. I. Bell, C. F. Burant, J. Takeda (подробнее…)
Как и для многих мембранных белков, трехмерная структура транспортера глюкозы неизвестна, поэтому молекулярный механизм транспорта глюкозы остается открытым вопросом. Однако кинетические исследования показывают, что переносчик глюкозы функционирует путем чередования двух конформационных состояний (1). В первой конформации сайт связывания глюкозы обращен к внешней стороне клетки. Связывание глюкозы с этим внешним сайтом вызывает конформационные изменения в транспортер, так что сайт связывания глюкозы теперь обращен внутрь клетки. Затем глюкоза может высвобождаться в цитозоль, после чего транспортер возвращается к своей исходной конформации.
Затем глюкоза может высвобождаться в цитозоль, после чего транспортер возвращается к своей исходной конформации.
Рисунок 12.17
Модель облегченной диффузии глюкозы. Транспортер глюкозы чередуется между двумя конформациями, в которых сайт связывания глюкозы попеременно экспонируется снаружи и внутри клетки. В первой показанной конформации (A) глюкоза (подробнее…)
Большинство клеток, включая эритроциты, подвергаются воздействию внеклеточных концентраций глюкозы, которые выше, чем концентрации внутри клетки, поэтому облегченная диффузия приводит к чистому транспорту глюкозы внутрь . Как только глюкоза поглощается этими клетками, она быстро метаболизируется, поэтому внутриклеточные концентрации глюкозы остаются низкими, и глюкоза продолжает транспортироваться в клетку из внеклеточной жидкости. Однако, поскольку конформационные изменения переносчика глюкозы обратимы, глюкоза может транспортироваться в противоположном направлении, просто обращая шаги в . Такой обратный поток имеет место, например, в клетках печени, в которых синтезируется глюкоза и высвобождается в кровоток.
Ионные каналы
В отличие от белков-носителей, канальные белки просто образуют открытые поры в мембране, позволяя небольшим молекулам соответствующего размера и заряда свободно проходить через липидный бислой. Одна группа белков каналов, обсуждавшаяся ранее, — это порины, которые обеспечивают свободное прохождение ионов и небольших полярных молекул через наружные мембраны бактерий (см. ). Канальные белки также обеспечивают прохождение молекул между клетками, соединенными щелевыми контактами, которые обсуждаются далее в этой главе. Плазматические мембраны многих клеток также содержат белки водных каналов (аквапорины), через которые молекулы воды способны пересекать мембрану гораздо быстрее, чем они могут диффундировать через бислой фосфолипидов. Однако наиболее хорошо охарактеризованными белками каналов являются ионные каналы, которые опосредуют прохождение ионов через плазматические мембраны. Хотя ионные каналы присутствуют в мембранах всех клеток, они особенно хорошо изучены в нервах и мышцах, где их регулируемое открытие и закрытие отвечает за передачу электрических сигналов.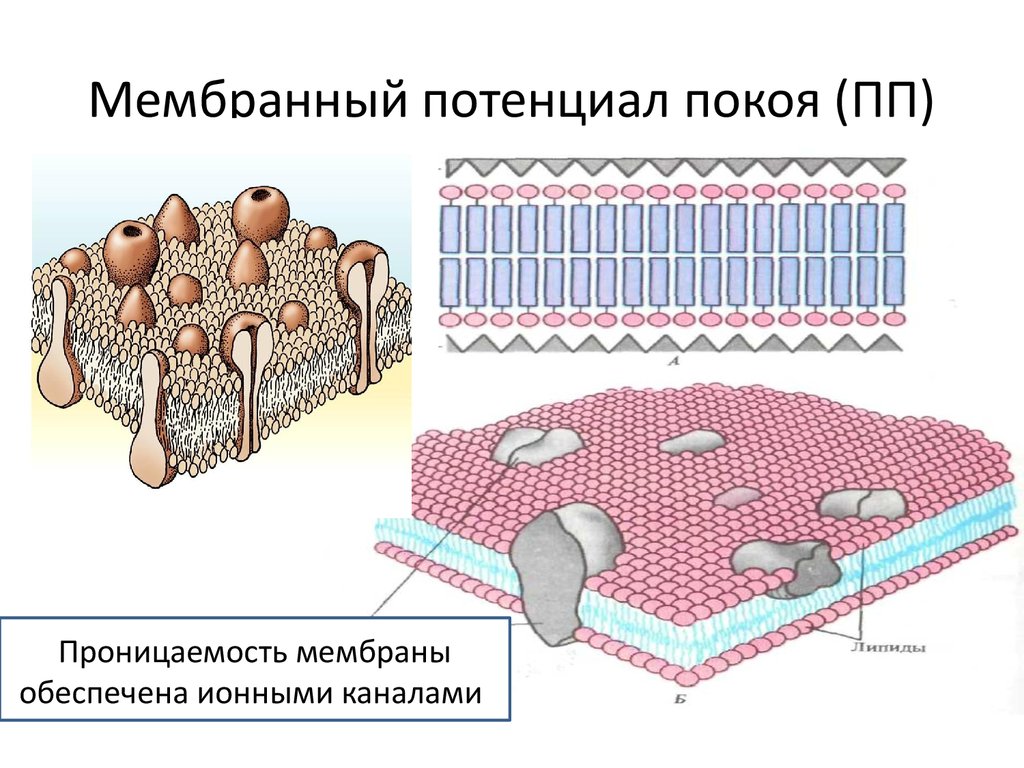
Три свойства ионных каналов лежат в основе их функции (). Во-первых, транспортировка по каналам чрезвычайно быстра. Более миллиона ионов в секунду проходят через открытые каналы — скорость потока примерно в тысячу раз превышает скорость транспорта белков-переносчиков. Во-вторых, ионные каналы обладают высокой селективностью, поскольку узкие поры в канале ограничивают прохождение ионов соответствующего размера и заряда. Таким образом, специфические белки каналов обеспечивают прохождение Na + , K + , Ca 2+ и Cl – через мембрану. В-третьих, большинство ионных каналов открыты не постоянно. Вместо этого открытие ионных каналов регулируется «воротами», которые кратковременно открываются в ответ на определенные раздражители. Некоторые каналы (называемые лиганд-зависимыми каналами ) открываются в ответ на связывание нейротрансмиттеров или других сигнальных молекул; другие ( потенциалзависимых канала ) открываются в ответ на изменения электрического потенциала на плазматической мембране.
Рисунок 12.18
Модель ионного канала. В закрытой конформации поток ионов блокируется воротами. Открытие ворот позволяет ионам быстро проходить через канал. Канал содержит узкую пору, которая ограничивает прохождение ионов соответствующего размера (подробнее…)
Фундаментальная роль ионных каналов в передаче электрических импульсов была выяснена в серии элегантных экспериментов, описанных Аланом Ходжкиным и Эндрю Хаксли. в 1952 году. Эти исследователи использовали в качестве модели гигантские нервные клетки кальмара. Аксоны этих гигантских нейронов имеют диаметр около 1 мм, что позволяет вводить электроды и измерять изменения мембранного потенциала, происходящие при передаче нервных импульсов. Используя этот подход, Ходжкин и Хаксли продемонстрировали, что эти изменения мембранного потенциала являются результатом регулируемого открытия и закрытия Na + и К + каналы в плазматической мембране. Впоследствии стало возможным изучать активность отдельных ионных каналов с помощью методики patch-clamp , разработанной Erwin Neher и Bert Sakmann в 1976 г. (). В этом методе микропипетка с диаметром наконечника около 1 мкм используется для выделения небольшого участка мембраны, что позволяет анализировать поток ионов через один канал и значительно повышает точность, с которой можно определить активность ионных каналов. учился.
(). В этом методе микропипетка с диаметром наконечника около 1 мкм используется для выделения небольшого участка мембраны, что позволяет анализировать поток ионов через один канал и значительно повышает точность, с которой можно определить активность ионных каналов. учился.
Рисунок 12.19
Метод накладного зажима. На кончике микропипетки выделяют небольшой участок мембраны. Затем внутри пипетки можно применять стимулы, что позволяет измерить поведение захваченного канала. (Адаптировано из E. Neher and B. Sakmann, 1992. (подробнее…)
Поток ионов через мембранные каналы зависит от установления ионных градиентов через плазматическую мембрану. Все клетки, включая нервные и мышечные, содержат ионных насосов (обсуждается в следующем разделе), которые используют энергию, полученную в результате гидролиза АТФ, для активного транспорта ионов через плазматическую мембрану.В результате ионный состав цитоплазмы существенно отличается от внеклеточной жидкости ().Например, Na + активно выкачивается из клеток, а К + закачивается. Таким образом, в аксоне кальмара концентрация Na + примерно в 10 раз выше во внеклеточной жидкости, чем внутри клетки, тогда как концентрация К + примерно в 20 раз выше в цитозоле, чем в окружающей среде.
Таким образом, в аксоне кальмара концентрация Na + примерно в 10 раз выше во внеклеточной жидкости, чем внутри клетки, тогда как концентрация К + примерно в 20 раз выше в цитозоле, чем в окружающей среде.
Таблица 12.1
Концентрации внеклеточных и внутриклеточных ионов.
Поскольку ионы электрически заряжены, их перенос приводит к возникновению электрического градиента на плазматической мембране. В покоящихся аксонах кальмара существует электрический потенциал около 60 мВ через плазматическую мембрану, при этом внутренняя часть клетки отрицательная по отношению к внешней (). Этот электрический потенциал возникает как от ионных насосов, так и от потока ионов через каналы, открытые в плазматической мембране покоящейся клетки. Плазматическая мембрана аксонов покоящегося кальмара содержит открытые K + , поэтому он более проницаем для K + , чем для Na + или других ионов. Следовательно, поток К + вносит наибольший вклад в мембранный потенциал покоя.
Рис. 12.20
Ионные градиенты и мембранный потенциал покоя аксона гигантского кальмара. Показаны только концентрации Na + и K + , поскольку именно эти ионы участвуют в передаче нервных импульсов. Na + выкачивается из клетки, а K + закачивается, (подробнее…)
Как обсуждалось в главе 10, поток ионов через мембрану управляется как концентрацией, так и компонентами напряжения электрохимического градиента. Например, концентрация K + внутри аксона кальмара в 20 раз выше, чем во внеклеточной жидкости, что приводит к выходу K + из клетки. Однако, поскольку K + заряжен положительно, этот выход K + из клетки генерирует электрический потенциал через мембрану, при этом внутренняя часть клетки становится отрицательно заряженной. Этот мембранный потенциал противостоит продолжающемуся потоку K + из клетки, и система приближается к равновесному состоянию, при котором мембранный потенциал уравновешивает градиент концентрации К + .
Количественно связь между концентрацией ионов и мембранным потенциалом определяется уравнением Нернста:
где В – равновесный потенциал в вольтах, R – газовая постоянная, T – абсолютная температура, z – заряд иона, F — постоянная Фарадея, а C o и C i — концентрации иона снаружи и внутри клетки соответственно. Равновесный потенциал существует отдельно для каждого иона, а мембранный потенциал определяется потоком всех ионов, пересекающих плазматическую мембрану. Однако, поскольку покоящиеся аксоны кальмаров более проницаемы для K + , чем для Na + или других ионов (включая Cl – ), мембранный потенциал покоя (-60 мВ) близок к равновесному потенциалу, определяемому внутриклеточным и внеклеточный K + концентрации (-75 мВ).
Когда нервные импульсы ( потенциалы действия ) проходят по аксонам, мембрана деполяризуется (). Мембранный потенциал изменяется от -60 мВ до примерно +30 мВ менее чем за миллисекунду, после чего снова становится отрицательным и возвращается к своему значению покоя. Эти изменения являются результатом быстрого последовательного открытия и закрытия потенциалзависимых каналов Na + и K + . Относительно небольшие начальные изменения мембранного потенциала (от -60 до примерно -40 мВ) приводят к быстрому открытию Na + канала. Это позволяет Na + поступать в клетку под действием как градиента его концентрации, так и мембранного потенциала. Внезапное поступление Na + приводит к сильному изменению мембранного потенциала, который увеличивается почти до +30 мВ, приближаясь к равновесному потенциалу Na + примерно +50 мВ. В это время каналы Na + инактивируются, а потенциалзависимые каналы K + открываются, существенно повышая проницаемость мембраны для K + . Затем K + быстро вытекает из клетки под действием как мембранного потенциала, так и градиента концентрации K + , что приводит к быстрому снижению мембранного потенциала примерно до -75 мВ (равновесный потенциал K + ).
Эти изменения являются результатом быстрого последовательного открытия и закрытия потенциалзависимых каналов Na + и K + . Относительно небольшие начальные изменения мембранного потенциала (от -60 до примерно -40 мВ) приводят к быстрому открытию Na + канала. Это позволяет Na + поступать в клетку под действием как градиента его концентрации, так и мембранного потенциала. Внезапное поступление Na + приводит к сильному изменению мембранного потенциала, который увеличивается почти до +30 мВ, приближаясь к равновесному потенциалу Na + примерно +50 мВ. В это время каналы Na + инактивируются, а потенциалзависимые каналы K + открываются, существенно повышая проницаемость мембраны для K + . Затем K + быстро вытекает из клетки под действием как мембранного потенциала, так и градиента концентрации K + , что приводит к быстрому снижению мембранного потенциала примерно до -75 мВ (равновесный потенциал K + ). Затем потенциалзависимые каналы K + инактивируются, и мембранный потенциал возвращается к уровню покоя -60 мВ, определяемому потоком K + и других ионов через каналы, которые остаются открытыми в нестимулированных клетках.
Затем потенциалзависимые каналы K + инактивируются, и мембранный потенциал возвращается к уровню покоя -60 мВ, определяемому потоком K + и других ионов через каналы, которые остаются открытыми в нестимулированных клетках.
Рис. 12.21
Мембранный потенциал и ионные каналы во время потенциала действия. (A) Изменения мембранного потенциала в одной точке гигантского аксона кальмара после стимула. E Na и E K – равновесные потенциалы Na + и K + соответственно. (B) Мембранный потенциал (подробнее…)
Деполяризация соседних областей плазматической мембраны позволяет потенциалам действия проходить по длине аксонов нервных клеток в виде электрических сигналов, что приводит к быстрой передаче нервных импульсов на большие расстояния. Например, аксоны двигательных нейронов человека могут иметь длину более метра. Прибытие потенциалов действия на окончание большинства нейронов сигнализирует о высвобождении нейротрансмиттеров, таких как ацетилхолин, которые передают сигналы между клетками в синапсах. Нейротрансмиттеры, высвобождаемые из пресинаптических клеток, связываются с рецепторами на мембранах постсинаптических клеток, где они действуют, открывая управляемые лигандом ионные каналы. Одним из наиболее хорошо охарактеризованных таких каналов является ацетилхолиновый рецептор мышечных клеток. Связывание ацетилхолина открывает канал, проницаемый как для Na + и К +. Это обеспечивает быстрый приток Na + , который деполяризует мембрану мышечной клетки и запускает потенциал действия. Затем потенциал действия приводит к открытию потенциалзависимых каналов Ca 2+ , что приводит к увеличению внутриклеточного количества Ca 2+ , что сигнализирует о сокращении (see ).
Нейротрансмиттеры, высвобождаемые из пресинаптических клеток, связываются с рецепторами на мембранах постсинаптических клеток, где они действуют, открывая управляемые лигандом ионные каналы. Одним из наиболее хорошо охарактеризованных таких каналов является ацетилхолиновый рецептор мышечных клеток. Связывание ацетилхолина открывает канал, проницаемый как для Na + и К +. Это обеспечивает быстрый приток Na + , который деполяризует мембрану мышечной клетки и запускает потенциал действия. Затем потенциал действия приводит к открытию потенциалзависимых каналов Ca 2+ , что приводит к увеличению внутриклеточного количества Ca 2+ , что сигнализирует о сокращении (see ).
Рисунок 12.22
Передача сигналов за счет высвобождения нейротрансмиттера в синапсе. Прибытие нервного импульса на окончание нейрона сигнализирует о слиянии синаптических пузырьков с плазматической мембраной, что приводит к высвобождению нейротрансмиттера из пресинаптической клетки в (далее. ..)
..)
Ацетилхолиновый рецептор, первоначально выделенный из электрического органа скатов Torpedo в 1970-х годах, является прототипом лиганд-управляемых каналов. Рецептор состоит из пяти субъединиц, расположенных в виде цилиндра в мембране (). Считается, что в закрытом состоянии поры канала блокируются боковыми цепями гидрофобных аминокислот. Связывание ацетилхолина вызывает конформационные изменения в рецепторе, так что эти гидрофобные боковые цепи смещаются из канала, открывая пору, через которую проходят положительно заряженные ионы, включая Na + и К +. Однако канал остается непроницаемым для отрицательно заряженных ионов, таких как Cl –, поскольку он выстлан отрицательно заряженными аминокислотами.
Рисунок 12.23
Модель рецептора ацетилхолина. Рецептор состоит из пяти субъединиц, расположенных вокруг центральной поры. Связывание ацетилхолина с участком во внеклеточной области рецептора вызывает аллостерические изменения, которые открывают ворота канала. Канал (подробнее…)
Канал (подробнее…)
Большую степень ионной селективности проявляют потенциалзависимые каналы Na + и K + . Каналы Na + более чем в десять раз более проницаемы для Na + , чем для K + , тогда как каналы K + более чем в тысячу раз более проницаемы для K + , чем для Na + . Селективность канала Na + может быть объяснена, по крайней мере, частично, на основе узких пор, которые действуют как размерный фильтр. Ионный радиус Na + (0,95 Å) меньше, чем у K + (1,33 Å), и считается, что пора канала Na + достаточно узкая, чтобы препятствовать прохождению ионов K + или более крупных ( ).
Рисунок 12.24
Ионная селективность каналов Na + . Узкая пора позволяет проходить Na + , связанным с одной молекулой воды, но препятствует прохождению ионов K + или более крупных ионов.
K + каналы также имеют узкие поры, препятствующие прохождению более крупных ионов. Однако, поскольку Na + имеет меньший ионный радиус, это не объясняет избирательной проницаемости этих каналов для K + . Селективность канала K + основана на другом механизме, который был выяснен при определении трехмерной структуры канала K + с помощью рентгеновской кристаллографии в 1998 г. (). Пора канала содержит фильтр с узкой селективностью, выстланный атомами карбонильного кислорода (C=O) из основной цепи полипептида. Когда К 9Ион 0061 + поступает в фильтр селективности, взаимодействия с этими карбонильными атомами кислорода вытесняют молекулы воды, с которыми связан K + , позволяя обезвоженному K + проходить через поры. Напротив, обезвоженный Na + слишком мал, чтобы взаимодействовать с этими карбонильными атомами кислорода в селективном фильтре, который удерживается жестко открытым. Следовательно, Na + остается связанным с молекулами воды в гидратированном комплексе, который слишком велик, чтобы пройти через канал.
Однако, поскольку Na + имеет меньший ионный радиус, это не объясняет избирательной проницаемости этих каналов для K + . Селективность канала K + основана на другом механизме, который был выяснен при определении трехмерной структуры канала K + с помощью рентгеновской кристаллографии в 1998 г. (). Пора канала содержит фильтр с узкой селективностью, выстланный атомами карбонильного кислорода (C=O) из основной цепи полипептида. Когда К 9Ион 0061 + поступает в фильтр селективности, взаимодействия с этими карбонильными атомами кислорода вытесняют молекулы воды, с которыми связан K + , позволяя обезвоженному K + проходить через поры. Напротив, обезвоженный Na + слишком мал, чтобы взаимодействовать с этими карбонильными атомами кислорода в селективном фильтре, который удерживается жестко открытым. Следовательно, Na + остается связанным с молекулами воды в гидратированном комплексе, который слишком велик, чтобы пройти через канал.
Рисунок 12.25
Избирательность каналов K + . Канал К + содержит фильтр узкой селективности, выстланный карбонильными атомами кислорода (С=О). Поры достаточно широки, чтобы обеспечить прохождение обезвоженного K + , из которого все связанные молекулы воды были вытеснены в виде (далее…)
Потенциалзависимых Na + , K + и Ca 2+ канала принадлежат к большому семейству родственных белков (2). Например, последовательность генома C. elegans выявил почти 200 генов, кодирующих ионные каналы, которые, по-видимому, необходимы, чтобы играть различные роли в клеточной передаче сигналов. Каналы K + состоят из четырех идентичных субъединиц, каждая из которых содержит две или шесть трансмембранных α-спиралей. Каналы Na + и Ca 2+ состоят из одной полипептидной цепи, но каждый полипептид содержит четыре повторяющихся домена, соответствующих субъединицам каналов K + . Запирание напряжения опосредовано одной из трансмембранных α-спиралей, которая содержит несколько положительно заряженных аминокислот. Мембранная деполяризация индуцирует перемещение этих положительных зарядов наружу клетки, сдвигая положение этого трансмембранного сегмента и открывая канал. Быстрая инактивация Na + и К + каналы при распространении потенциалов действия затем опосредованы цитоплазматическими участками полипептидной цепи, которые связываются с цитоплазматическим устьем поры канала и препятствуют дальнейшему току ионов ().
Запирание напряжения опосредовано одной из трансмембранных α-спиралей, которая содержит несколько положительно заряженных аминокислот. Мембранная деполяризация индуцирует перемещение этих положительных зарядов наружу клетки, сдвигая положение этого трансмембранного сегмента и открывая канал. Быстрая инактивация Na + и К + каналы при распространении потенциалов действия затем опосредованы цитоплазматическими участками полипептидной цепи, которые связываются с цитоплазматическим устьем поры канала и препятствуют дальнейшему току ионов ().
Рисунок 12.26
Структуры потенциалзависимых катионных каналов. Каналы K + , Na + и Ca 2+ принадлежат к семейству родственных белков. Канал К + образован объединением четырех идентичных субъединиц, одна из которых показана. На 9Канал 0061 + состоит из одного полипептида (подробнее…)
Рисунок 12.27
Инактивация каналов K + и Na + . После потенциалзависимого открытия каналы K + и Na + быстро инактивируются за счет связывания цитоплазматических частей полипептидных цепей с порой. Для канала K + инактивация опосредована шаровой цепью (подробнее…)
После потенциалзависимого открытия каналы K + и Na + быстро инактивируются за счет связывания цитоплазматических частей полипептидных цепей с порой. Для канала K + инактивация опосредована шаровой цепью (подробнее…)
Широкий спектр ионных каналов (включая Ca 2+ и Cl – каналы) реагируют на разные нейротрансмиттеры или открываются и закрываются с разной кинетикой после деполяризации мембраны. Согласованные действия этих многочисленных каналов ответственны за сложность передачи сигналов в нервной системе. Более того, как будет показано в следующей главе, роль ионных каналов не ограничивается электрически возбудимыми нервными и мышечными клетками; они также играют важную роль в передаче сигналов в других типах клеток. Таким образом, регулируемое открытие и закрытие ионных каналов обеспечивает клетки чувствительным и универсальным механизмом реагирования на различные стимулы окружающей среды.
Активный транспорт, управляемый гидролизом АТФ
Чистый поток молекул за счет облегченной диффузии либо через белки-носители, либо через белки-каналы всегда энергетически направлен вниз в направлении, определяемом электрохимическими градиентами через мембрану. Однако во многих случаях клетка должна транспортировать молекулы против градиента их концентрации. При активном транспорте энергия, обеспечиваемая другой связанной реакцией (например, гидролизом АТФ), используется для перемещения молекул в гору в энергетически невыгодном направлении.
Однако во многих случаях клетка должна транспортировать молекулы против градиента их концентрации. При активном транспорте энергия, обеспечиваемая другой связанной реакцией (например, гидролизом АТФ), используется для перемещения молекул в гору в энергетически невыгодном направлении.
Ионные насосы, отвечающие за поддержание градиента ионов через плазматическую мембрану, являются важными примерами активного транспорта, управляемого непосредственно гидролизом АТФ. Как обсуждалось ранее (см.), концентрация Na + примерно в десять раз выше снаружи, чем внутри клеток, тогда как концентрация K + выше внутри, чем снаружи. Эти градиенты ионов поддерживаются насосом Na + -K + (также называемая Na + -K + АТФаза ), которая использует энергию, полученную в результате гидролиза АТФ, для транспорта Na + и K + против их электрохимических градиентов. Этот процесс является результатом конформационных изменений в помпе, вызванных АТФ. Во-первых, ионы Na + связываются с центрами высокого сродства внутри клетки. Это связывание стимулирует гидролиз АТФ и фосфорилирование помпы, вызывая конформационные изменения, которые обнажают Na + – связывает сайты снаружи клетки и снижает их сродство к Na + . Следовательно, связанный Na + высвобождается во внеклеточную жидкость. В то же время на клеточной поверхности экспонируются высокоаффинные сайты связывания К + -. Связывание внеклеточного K + с этими сайтами затем стимулирует гидролиз фосфатной группы, связанной с насосом, что вызывает второе конформационное изменение, открывая сайты связывания K + цитозолю и снижая их аффинность связывания, так что K + высвобождается внутри клетки. Насос имеет три сайта связывания для Na + и два для K + , поэтому каждый цикл переносит через плазматическую мембрану три Na + и два K + за счет одной молекулы АТФ.
Этот процесс является результатом конформационных изменений в помпе, вызванных АТФ. Во-первых, ионы Na + связываются с центрами высокого сродства внутри клетки. Это связывание стимулирует гидролиз АТФ и фосфорилирование помпы, вызывая конформационные изменения, которые обнажают Na + – связывает сайты снаружи клетки и снижает их сродство к Na + . Следовательно, связанный Na + высвобождается во внеклеточную жидкость. В то же время на клеточной поверхности экспонируются высокоаффинные сайты связывания К + -. Связывание внеклеточного K + с этими сайтами затем стимулирует гидролиз фосфатной группы, связанной с насосом, что вызывает второе конформационное изменение, открывая сайты связывания K + цитозолю и снижая их аффинность связывания, так что K + высвобождается внутри клетки. Насос имеет три сайта связывания для Na + и два для K + , поэтому каждый цикл переносит через плазматическую мембрану три Na + и два K + за счет одной молекулы АТФ.
Рисунок 12.28
Модель для работы насоса Na + -K + .
Важность помпы Na + -K + подтверждается тем фактом, что она потребляет почти 25% АТФ, используемого многими животными клетками. Одна из важнейших ролей Na + и К + градиенты, установленные насосом, — это распространение электрических сигналов в нерве и мышце. Как будет сказано ниже, градиент Na + , создаваемый помпой, также используется для обеспечения активного транспорта множества других молекул. Еще одна важная роль насоса Na + -K + в большинстве клеток животных заключается в поддержании осмотического баланса и объема клетки. Цитоплазма содержит высокую концентрацию органических молекул, включая макромолекулы, аминокислоты, сахара и нуклеотиды. В отсутствие противовеса это будет стимулировать поток воды внутрь за счет осмоса, что, если его не остановить, приведет к набуханию и, в конечном итоге, к разрыву клетки. Необходимый противовес обеспечивается градиентами ионов, установленными Na + -К + насос (). В частности, насос устанавливает более высокую концентрацию Na + снаружи, чем внутри клетки. Как уже обсуждалось, поток K + через открытые каналы дополнительно устанавливает электрический потенциал через плазматическую мембрану. Этот мембранный потенциал, в свою очередь, выталкивает Cl – из клетки, поэтому концентрация Cl – (как и Na + ) примерно в десять раз выше во внеклеточной жидкости, чем в цитоплазме. Эти различия в концентрациях ионов уравновешивают высокие концентрации органических молекул внутри клеток, выравнивая осмотическое давление и предотвращая чистый приток воды.
Необходимый противовес обеспечивается градиентами ионов, установленными Na + -К + насос (). В частности, насос устанавливает более высокую концентрацию Na + снаружи, чем внутри клетки. Как уже обсуждалось, поток K + через открытые каналы дополнительно устанавливает электрический потенциал через плазматическую мембрану. Этот мембранный потенциал, в свою очередь, выталкивает Cl – из клетки, поэтому концентрация Cl – (как и Na + ) примерно в десять раз выше во внеклеточной жидкости, чем в цитоплазме. Эти различия в концентрациях ионов уравновешивают высокие концентрации органических молекул внутри клеток, выравнивая осмотическое давление и предотвращая чистый приток воды.
Рис. 12.29
Градиенты ионов через плазматическую мембрану типичной клетки млекопитающих. Концентрации Na + и Cl – выше снаружи, чем внутри клетки, тогда как концентрация К + выше внутри, чем снаружи.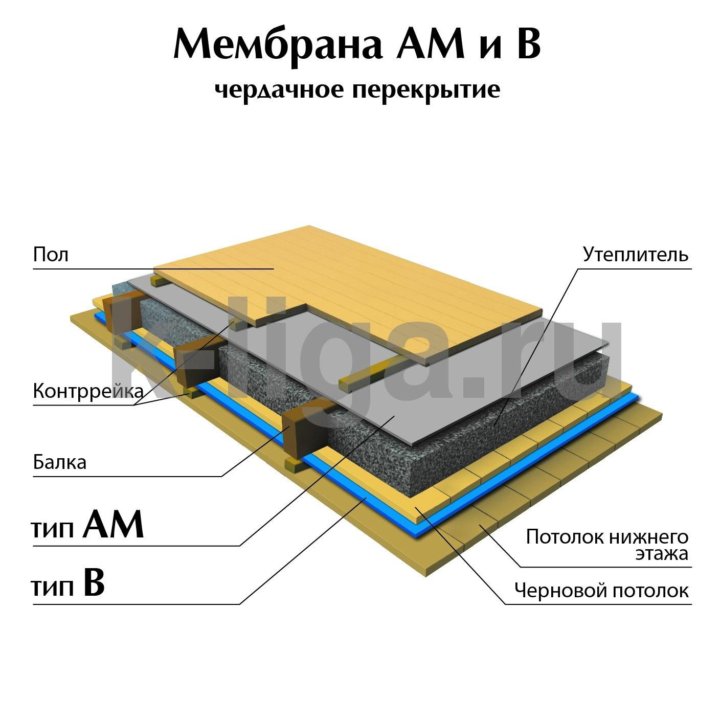 Низкие концентрации Na + и Cl – уравновешивают (подробнее…)
Низкие концентрации Na + и Cl – уравновешивают (подробнее…)
Активный транспорт Ca 2+ через плазматическую мембрану управляется насосом Ca 2+ , который структурно родственен до На + -K + насос и аналогичным образом питается от гидролиза АТФ. Насос Ca 2+ транспортирует Ca 2+ из клетки, поэтому внутриклеточные концентрации Ca 2+ чрезвычайно низки: приблизительно 0,1 мкМ, по сравнению с внеклеточными концентрациями около 1 м M . Эта низкая внутриклеточная концентрация Ca 2+ делает клетку чувствительной к небольшому увеличению внутриклеточного уровня Ca 2+ . Такое временное увеличение внутриклеточного Ca 2+ играют важную роль в клеточной передаче сигналов, как уже отмечалось в отношении мышечного сокращения (см. ) и обсуждается далее в следующей главе.
Аналогичные ионные насосы в плазматических мембранах бактерий, дрожжей и растительных клеток ответственны за активный транспорт H + из клетки.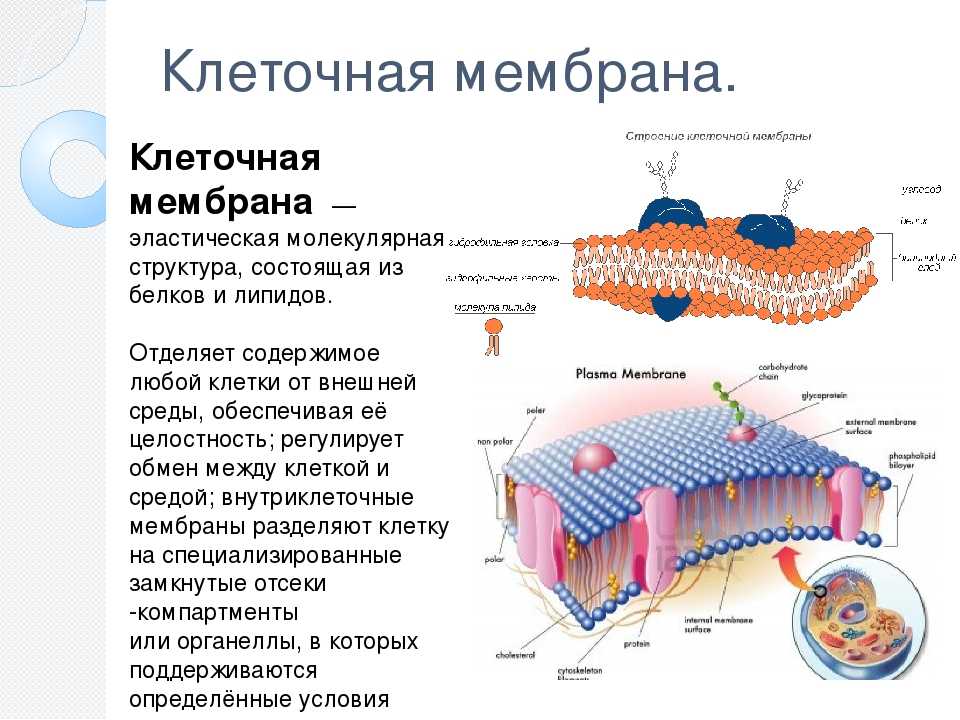 Кроме того, H + активно выкачиваются из клеток, выстилающих желудок, что приводит к повышению кислотности желудочного сока. Структурно различные помпы ответственны за активный транспорт H + на лизосомы и эндосомы (см.). Тем не менее, третий тип насоса H + иллюстрируется АТФ-синтазами митохондрий и хлоропластов: в этих случаях насосы можно рассматривать как работающие в обратном направлении, когда движение ионов по электрохимическому градиенту используется для управления синтезом АТФ.
Кроме того, H + активно выкачиваются из клеток, выстилающих желудок, что приводит к повышению кислотности желудочного сока. Структурно различные помпы ответственны за активный транспорт H + на лизосомы и эндосомы (см.). Тем не менее, третий тип насоса H + иллюстрируется АТФ-синтазами митохондрий и хлоропластов: в этих случаях насосы можно рассматривать как работающие в обратном направлении, когда движение ионов по электрохимическому градиенту используется для управления синтезом АТФ.
Самое большое семейство мембранных транспортеров состоит из транспортеров ABC, названных так потому, что они характеризуются высококонсервативными АТФ-связывающими доменами или A TP- b inding c активы (). Как в прокариотических, так и в эукариотических клетках идентифицировано более сотни членов семейства. У бактерий транспортеры ABC используют энергию, полученную в результате гидролиза АТФ, для транспорта широкого спектра молекул, включая ионы, сахара и аминокислоты. В эукариотических клетках был обнаружен первый переносчик ABC как продукт гена (называемого геном множественной лекарственной устойчивости, или mdr ), который делает раковые клетки устойчивыми к различным лекарствам, используемым в химиотерапии. В настоящее время идентифицированы два переносчика MDR. Обычно они экспрессируются в различных клетках, где они удаляют потенциально токсичные чужеродные соединения. Например, экспрессия переносчика MDR в капиллярных эндотелиальных клетках головного мозга, по-видимому, играет важную роль в защите мозга от токсичных химических веществ. К сожалению, транспортеры MDR часто экспрессируются на высоких уровнях в раковых клетках, где они распознают различные лекарства и выкачивают их из клеток, тем самым делая раковые клетки устойчивыми к широкому спектру химиотерапевтических агентов и создавая серьезное препятствие для успешного лечения рака. .
В эукариотических клетках был обнаружен первый переносчик ABC как продукт гена (называемого геном множественной лекарственной устойчивости, или mdr ), который делает раковые клетки устойчивыми к различным лекарствам, используемым в химиотерапии. В настоящее время идентифицированы два переносчика MDR. Обычно они экспрессируются в различных клетках, где они удаляют потенциально токсичные чужеродные соединения. Например, экспрессия переносчика MDR в капиллярных эндотелиальных клетках головного мозга, по-видимому, играет важную роль в защите мозга от токсичных химических веществ. К сожалению, транспортеры MDR часто экспрессируются на высоких уровнях в раковых клетках, где они распознают различные лекарства и выкачивают их из клеток, тем самым делая раковые клетки устойчивыми к широкому спектру химиотерапевтических агентов и создавая серьезное препятствие для успешного лечения рака. .
Рисунок 12.30
Структура транспортера ABC. Базовая структурная единица транспортеров ABC состоит из шести трансмембранных доменов, за которыми следует АТФ-связывающая кассета.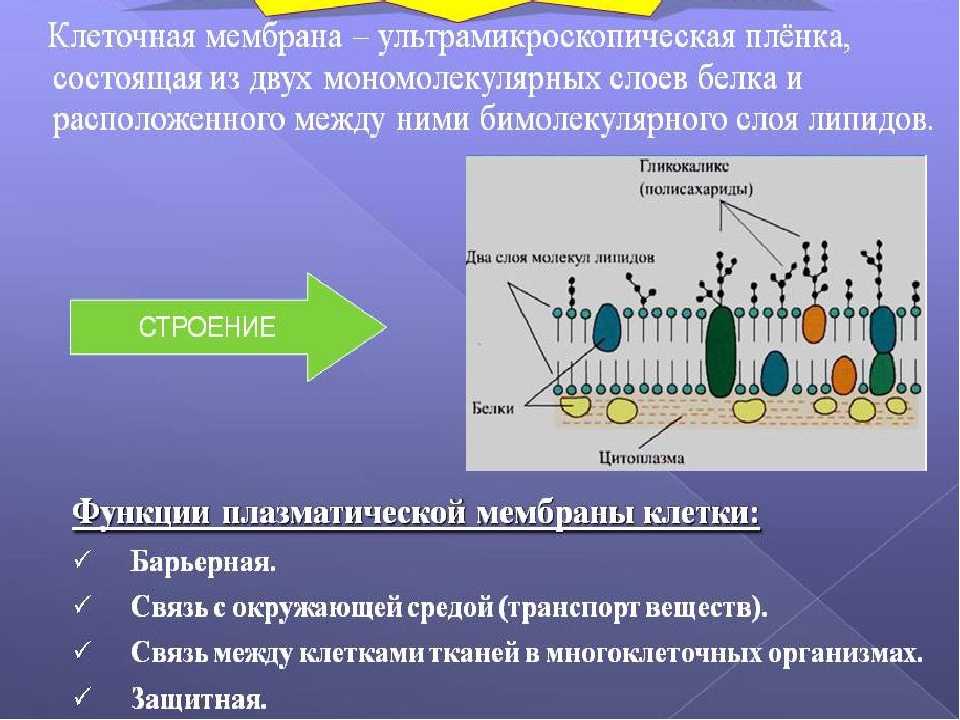 В транспортерах плазматической мембраны (таких как показанный транспортер MDR) две из этих единиц являются (подробнее…)
В транспортерах плазматической мембраны (таких как показанный транспортер MDR) две из этих единиц являются (подробнее…)
Еще одним важным с медицинской точки зрения членом семейства транспортеров ABC является ген, ответственный за муковисцидоз. Хотя он является членом семейства ABC, продукт этого гена (называемый регулятором трансмембранной проводимости при муковисцидозе, или CFTR) функционирует как Cl 9.0061- канал в эпителиальных клетках и дефектный транспорт Cl – характерны для заболевания. Канал CFTR Cl – также необычен тем, что для его открытия, по-видимому, требуется как гидролиз АТФ, так и зависимое от цАМФ фосфорилирование. Структурная основа функции CFTR как регулируемого ионного канала еще предстоит выяснить в ходе будущих исследований.
Активный транспорт, управляемый ионными градиентами
Ионные насосы и АВС-переносчики, рассмотренные в предыдущем разделе, используют энергию, полученную непосредственно в результате гидролиза АТФ, для транспортировки молекул против их электрохимических градиентов.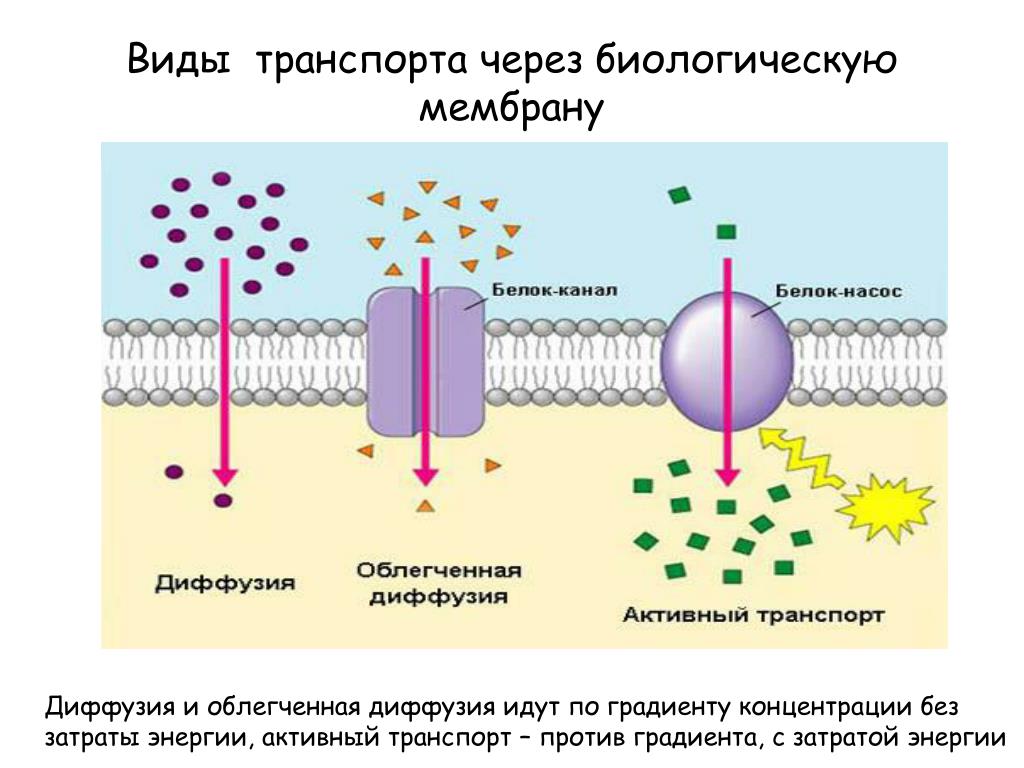 Другие молекулы транспортируются против градиента их концентрации с использованием энергии, полученной не от гидролиза АТФ, а от связанного транспорта второй молекулы в энергетически выгодном направлении. На 9Градиент 0061 + , созданный насосом Na + -K + , обеспечивает источник энергии, который часто используется для обеспечения активного транспорта сахаров, аминокислот и ионов в клетках млекопитающих. Аналогичную роль играют градиенты H + , установленные насосами H + клеток бактерий, дрожжей и растений.
Другие молекулы транспортируются против градиента их концентрации с использованием энергии, полученной не от гидролиза АТФ, а от связанного транспорта второй молекулы в энергетически выгодном направлении. На 9Градиент 0061 + , созданный насосом Na + -K + , обеспечивает источник энергии, который часто используется для обеспечения активного транспорта сахаров, аминокислот и ионов в клетках млекопитающих. Аналогичную роль играют градиенты H + , установленные насосами H + клеток бактерий, дрожжей и растений.
Эпителиальные клетки, выстилающие кишечник, представляют собой хороший пример активного транспорта, управляемого градиентом Na + . Эти клетки используют активные транспортные системы в апикальных областях своих плазматических мембран для захвата пищевых сахаров и аминокислот из просвета кишечника. Поглощение глюкозы, например, осуществляется транспортером, который координированно транспортирует два Na + и одна глюкоза в клетку ().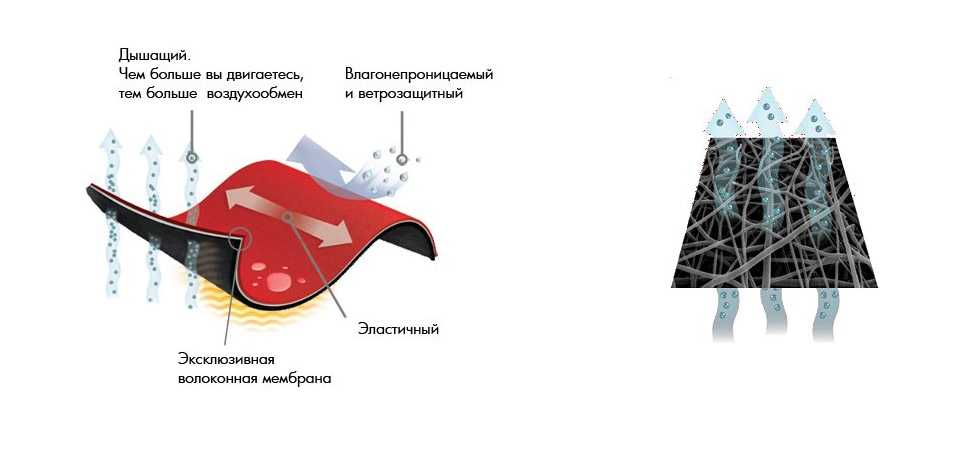 Поток Na + по его электрохимическому градиенту обеспечивает энергию, необходимую для поглощения пищевой глюкозы и накопления высоких концентраций внутриклеточной глюкозы. Затем глюкоза высвобождается в подлежащую соединительную ткань (которая содержит кровеносные капилляры) на базолатеральной поверхности кишечного эпителия, где она транспортируется по градиенту концентрации за счет облегченной диффузии (12). Таким образом, поглощение глюкозы из просвета кишечника и ее высвобождение в кровоток является хорошим примером поляризованной функции эпителиальных клеток, которая является результатом специфической локализации активного транспорта и облегченной диффузии переносчиков в апикальный и базолатеральный домены плазматической мембраны. , соответственно.
Поток Na + по его электрохимическому градиенту обеспечивает энергию, необходимую для поглощения пищевой глюкозы и накопления высоких концентраций внутриклеточной глюкозы. Затем глюкоза высвобождается в подлежащую соединительную ткань (которая содержит кровеносные капилляры) на базолатеральной поверхности кишечного эпителия, где она транспортируется по градиенту концентрации за счет облегченной диффузии (12). Таким образом, поглощение глюкозы из просвета кишечника и ее высвобождение в кровоток является хорошим примером поляризованной функции эпителиальных клеток, которая является результатом специфической локализации активного транспорта и облегченной диффузии переносчиков в апикальный и базолатеральный домены плазматической мембраны. , соответственно.
Рис. 12.31
Активный транспорт глюкозы. Активный транспорт, управляемый градиентом Na + , отвечает за поглощение глюкозы из просвета кишечника. Транспортер координированно связывает и транспортирует в клетку одну глюкозу и две Na + .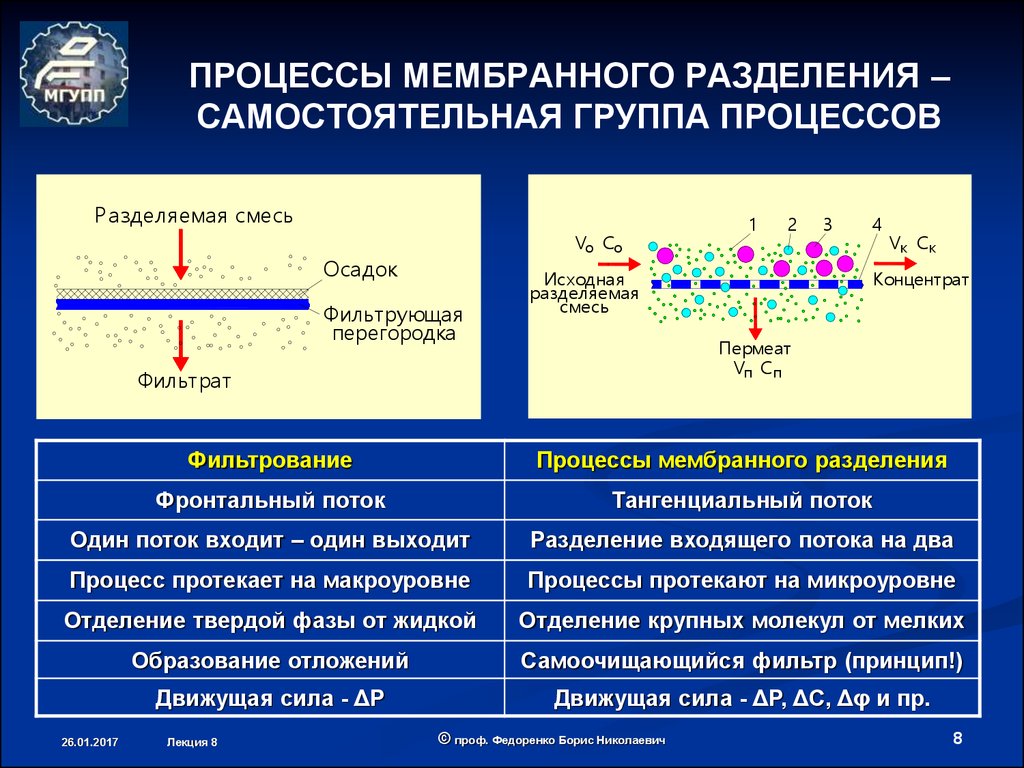 Транспорт Na + (подробнее…)
Транспорт Na + (подробнее…)
Рис. 12.32
Транспорт глюкозы эпителиальными клетками кишечника. Транспортер в апикальном домене плазматической мембраны отвечает за активное поглощение глюкозы (путем котранспорта с Na + ) из просвета кишечника. В результате пищевая глюкоза поглощается и (подробнее…)
Координированное поглощение глюкозы и Na + является примером симпорта, транспорта двух молекул в одном направлении. Напротив, облегченная диффузия глюкозы является примером унипорта, транспорта только одной молекулы. Активный транспорт также может осуществляться антипортом, при котором две молекулы транспортируются в противоположных направлениях (1). Например, Са 2+ экспортируется из клеток не только насосом Ca 2+ , но и антипортером Na + -Ca 2+ , который транспортирует Na + в клетку и Ca 2+ наружу. Другим примером является обменный белок Na + -H + , который участвует в регуляции внутриклеточного pH. Антипортер Na + -H + связывает транспорт Na + в клетку с экспортом H + , тем самым удаляя избыток H + , образующиеся в результате метаболических реакций и препятствующие закислению цитоплазмы.
Антипортер Na + -H + связывает транспорт Na + в клетку с экспортом H + , тем самым удаляя избыток H + , образующиеся в результате метаболических реакций и препятствующие закислению цитоплазмы.
Рисунок 12.33
Примеры антипорта. Ca 2+ и H + экспортируются из клеток антипортерами, которые связывают их экспорт с энергетически выгодным импортом Na + .
Коробка
Молекулярная медицина: кистозный фиброз.
Транспорт малых молекул — клетка
Внутренний состав клетки сохраняется, поскольку плазматическая мембрана избирательно проницаема для малых молекул. Большинство биологических молекул не способны диффундировать через фосфолипидный бислой, поэтому плазматическая мембрана образует барьер, блокирующий свободный обмен молекул между цитоплазмой и внешней средой клетки. Специфические транспортные белки (белки-носители и белковые каналы) опосредуют избирательное прохождение малых молекул через мембрану, позволяя клетке контролировать состав своей цитоплазмы.
Пассивная диффузия
Простейший механизм пересечения молекулами плазматической мембраны — пассивная диффузия. Во время пассивной диффузии молекула просто растворяется в бислое фосфолипидов, диффундирует через него, а затем растворяется в водном растворе на другой стороне мембраны. Никакие мембранные белки не участвуют, и направление транспорта определяется просто относительными концентрациями молекулы внутри и снаружи клетки. Чистый поток молекул всегда направлен вниз по градиенту их концентрации — из отсека с высокой концентрацией в отсек с более низкой концентрацией молекулы.
Таким образом, пассивная диффузия представляет собой неселективный процесс, при котором любая молекула, способная растворяться в двойном слое фосфолипидов, способна пересекать плазматическую мембрану и уравновешиваться между внутренней и внешней частью клетки. Важно отметить, что только небольшие, относительно гидрофобные молекулы способны диффундировать через бислой фосфолипидов со значительной скоростью.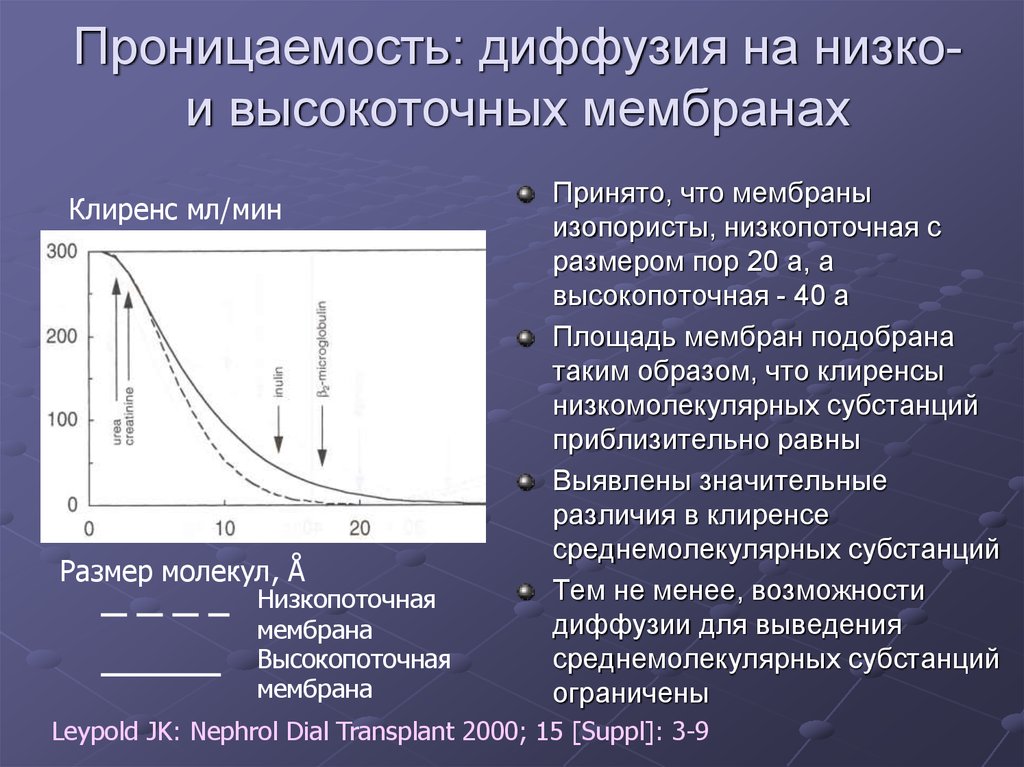 Так, газы (такие как O 2 и CO 2 ), гидрофобные молекулы (такие как бензол) и небольшие полярные, но незаряженные молекулы (такие как H 2 O и этанол) способны диффундировать через плазматическую мембрану. Однако другие биологические молекулы не способны растворяться в гидрофобной внутренней части двойного слоя фосфолипидов. Следовательно, более крупные незаряженные полярные молекулы, такие как глюкоза, не могут пересечь плазматическую мембрану путем пассивной диффузии, как и заряженные молекулы любого размера (включая небольшие ионы, такие как H + , Na + , K + и Cl). – ). Прохождение этих молекул через мембрану вместо этого требует активности специфических транспортных и канальных белков, которые, таким образом, контролируют движение большинства биологических молекул в клетку и из нее.
Так, газы (такие как O 2 и CO 2 ), гидрофобные молекулы (такие как бензол) и небольшие полярные, но незаряженные молекулы (такие как H 2 O и этанол) способны диффундировать через плазматическую мембрану. Однако другие биологические молекулы не способны растворяться в гидрофобной внутренней части двойного слоя фосфолипидов. Следовательно, более крупные незаряженные полярные молекулы, такие как глюкоза, не могут пересечь плазматическую мембрану путем пассивной диффузии, как и заряженные молекулы любого размера (включая небольшие ионы, такие как H + , Na + , K + и Cl). – ). Прохождение этих молекул через мембрану вместо этого требует активности специфических транспортных и канальных белков, которые, таким образом, контролируют движение большинства биологических молекул в клетку и из нее.
Рисунок 12.15
Проницаемость фосфолипидных бислоев. Газы, гидрофобные молекулы и небольшие полярные незаряженные молекулы могут диффундировать через бислои фосфолипидов..jpg) Более крупные полярные молекулы и заряженные молекулы не могут.
Более крупные полярные молекулы и заряженные молекулы не могут.
Облегченная диффузия и белки-носители
Облегченная диффузия, как и пассивная, включает движение молекул в направлении, определяемом их относительными концентрациями внутри и снаружи клетки. Никакого внешнего источника энергии не предусмотрено, поэтому молекулы движутся через мембрану в направлении, определяемом их градиентами концентрации и, в случае заряженных молекул, электрическим потенциалом на мембране. Однако облегченная диффузия отличается от пассивной диффузии тем, что транспортируемые молекулы не растворяются в двойном слое фосфолипидов. Вместо этого их прохождение опосредуется белками, которые позволяют транспортируемым молекулам пересекать мембрану, не взаимодействуя непосредственно с ее гидрофобной внутренней частью. Таким образом, облегченная диффузия позволяет полярным и заряженным молекулам, таким как углеводы, аминокислоты, нуклеозиды и ионы, пересекать плазматическую мембрану.
Обычно различают два класса белков, которые опосредуют облегченную диффузию: белки-носители и белки-каналы. Белки-переносчики связывают определенные молекулы для транспортировки на одной стороне мембраны. Затем они претерпевают конформационные изменения, которые позволяют молекуле пройти через мембрану и выйти на другую сторону. Напротив, канальные белки (см. следующий раздел) образуют открытые поры через мембрану, обеспечивая свободную диффузию любой молекулы соответствующего размера и заряда.
Белки-переносчики связывают определенные молекулы для транспортировки на одной стороне мембраны. Затем они претерпевают конформационные изменения, которые позволяют молекуле пройти через мембрану и выйти на другую сторону. Напротив, канальные белки (см. следующий раздел) образуют открытые поры через мембрану, обеспечивая свободную диффузию любой молекулы соответствующего размера и заряда.
Белки-носители отвечают за облегченную диффузию сахаров, аминокислот и нуклеозидов через плазматические мембраны большинства клеток. Поглощение глюкозы, которая служит основным источником метаболической энергии, является одной из наиболее важных транспортных функций плазматической мембраны, а транспортер глюкозы является хорошо изученным примером белка-переносчика. Транспортер глюкозы первоначально был идентифицирован как белок массой 55 кД в эритроцитах человека, в которых он составляет примерно 5% от общего белка мембраны. Последующее выделение и анализ последовательности клона кДНК выявили, что переносчик глюкозы имеет 12 α-спиральных трансмембранных сегментов — структура, типичная для многих белков-переносчиков (12). Эти трансмембранные α-спирали содержат преимущественно гидрофобные аминокислоты, но некоторые из них также содержат полярные аминокислотные остатки, которые, как считается, образуют сайт связывания глюкозы внутри белка.
Эти трансмембранные α-спирали содержат преимущественно гидрофобные аминокислоты, но некоторые из них также содержат полярные аминокислотные остатки, которые, как считается, образуют сайт связывания глюкозы внутри белка.
Рисунок 12.16
Структура переносчика глюкозы. Транспортер глюкозы имеет 12 трансмембранных α-спиралей. Полярные аминокислотные остатки, расположенные внутри фосфолипидного бислоя, обозначены темно-фиолетовыми кружками. (Адаптировано из G. I. Bell, C. F. Burant, J. Takeda (подробнее…)
Как и для многих мембранных белков, трехмерная структура транспортера глюкозы неизвестна, поэтому молекулярный механизм транспорта глюкозы остается открытым вопросом. Однако кинетические исследования показывают, что переносчик глюкозы функционирует путем чередования двух конформационных состояний (1). В первой конформации сайт связывания глюкозы обращен к внешней стороне клетки. Связывание глюкозы с этим внешним сайтом вызывает конформационные изменения в транспортер, так что сайт связывания глюкозы теперь обращен внутрь клетки. Затем глюкоза может высвобождаться в цитозоль, после чего транспортер возвращается к своей исходной конформации.
Затем глюкоза может высвобождаться в цитозоль, после чего транспортер возвращается к своей исходной конформации.
Рисунок 12.17
Модель облегченной диффузии глюкозы. Транспортер глюкозы чередуется между двумя конформациями, в которых сайт связывания глюкозы попеременно экспонируется снаружи и внутри клетки. В первой показанной конформации (A) глюкоза (подробнее…)
Большинство клеток, включая эритроциты, подвергаются воздействию внеклеточных концентраций глюкозы, которые выше, чем концентрации внутри клетки, поэтому облегченная диффузия приводит к чистому транспорту глюкозы внутрь . Как только глюкоза поглощается этими клетками, она быстро метаболизируется, поэтому внутриклеточные концентрации глюкозы остаются низкими, и глюкоза продолжает транспортироваться в клетку из внеклеточной жидкости. Однако, поскольку конформационные изменения переносчика глюкозы обратимы, глюкоза может транспортироваться в противоположном направлении, просто обращая шаги в . Такой обратный поток имеет место, например, в клетках печени, в которых синтезируется глюкоза и высвобождается в кровоток.
Ионные каналы
В отличие от белков-носителей, канальные белки просто образуют открытые поры в мембране, позволяя небольшим молекулам соответствующего размера и заряда свободно проходить через липидный бислой. Одна группа белков каналов, обсуждавшаяся ранее, — это порины, которые обеспечивают свободное прохождение ионов и небольших полярных молекул через наружные мембраны бактерий (см. ). Канальные белки также обеспечивают прохождение молекул между клетками, соединенными щелевыми контактами, которые обсуждаются далее в этой главе. Плазматические мембраны многих клеток также содержат белки водных каналов (аквапорины), через которые молекулы воды способны пересекать мембрану гораздо быстрее, чем они могут диффундировать через бислой фосфолипидов. Однако наиболее хорошо охарактеризованными белками каналов являются ионные каналы, которые опосредуют прохождение ионов через плазматические мембраны. Хотя ионные каналы присутствуют в мембранах всех клеток, они особенно хорошо изучены в нервах и мышцах, где их регулируемое открытие и закрытие отвечает за передачу электрических сигналов.
Три свойства ионных каналов лежат в основе их функции (). Во-первых, транспортировка по каналам чрезвычайно быстра. Более миллиона ионов в секунду проходят через открытые каналы — скорость потока примерно в тысячу раз превышает скорость транспорта белков-переносчиков. Во-вторых, ионные каналы обладают высокой селективностью, поскольку узкие поры в канале ограничивают прохождение ионов соответствующего размера и заряда. Таким образом, специфические белки каналов обеспечивают прохождение Na + , K + , Ca 2+ и Cl – через мембрану. В-третьих, большинство ионных каналов открыты не постоянно. Вместо этого открытие ионных каналов регулируется «воротами», которые кратковременно открываются в ответ на определенные раздражители. Некоторые каналы (называемые лиганд-зависимыми каналами ) открываются в ответ на связывание нейротрансмиттеров или других сигнальных молекул; другие ( потенциалзависимых канала ) открываются в ответ на изменения электрического потенциала на плазматической мембране.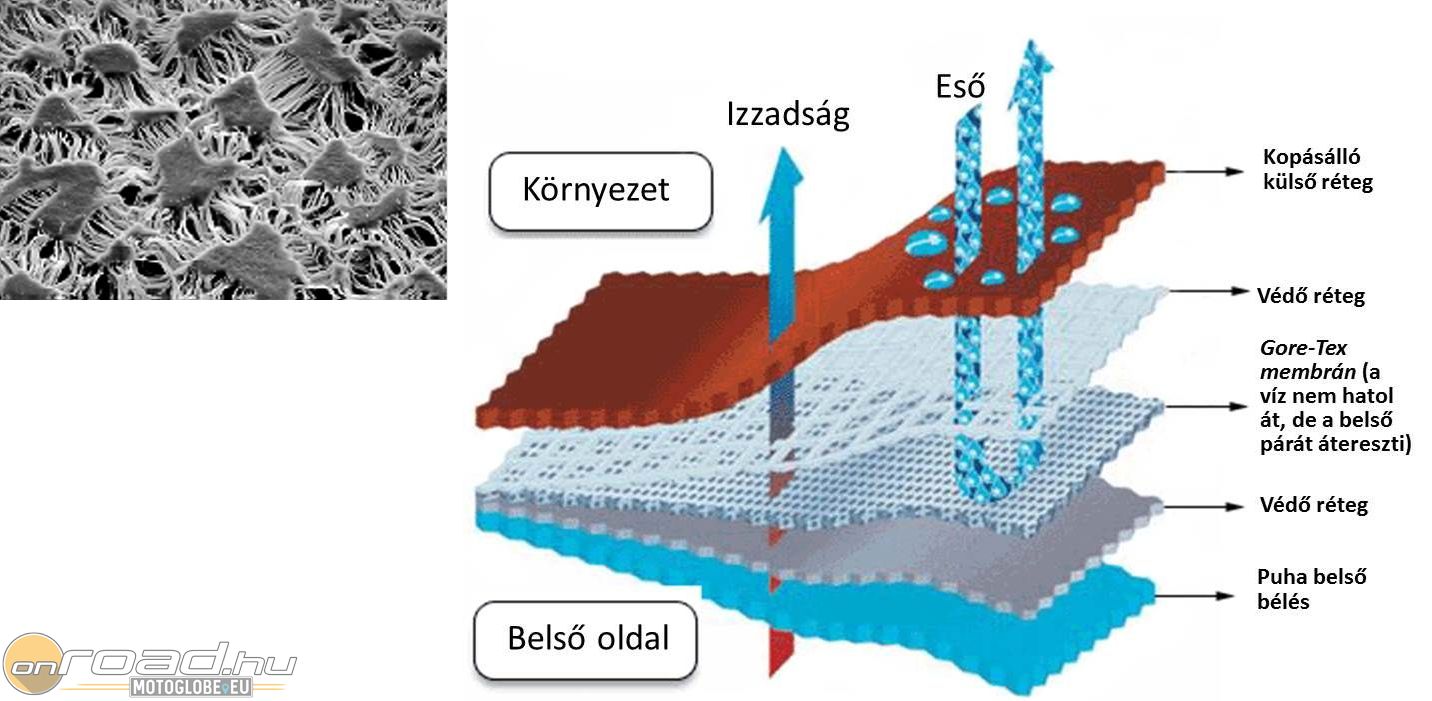
Рисунок 12.18
Модель ионного канала. В закрытой конформации поток ионов блокируется воротами. Открытие ворот позволяет ионам быстро проходить через канал. Канал содержит узкую пору, которая ограничивает прохождение ионов соответствующего размера (подробнее…)
Фундаментальная роль ионных каналов в передаче электрических импульсов была выяснена в серии элегантных экспериментов, описанных Аланом Ходжкиным и Эндрю Хаксли. в 1952 году. Эти исследователи использовали в качестве модели гигантские нервные клетки кальмара. Аксоны этих гигантских нейронов имеют диаметр около 1 мм, что позволяет вводить электроды и измерять изменения мембранного потенциала, происходящие при передаче нервных импульсов. Используя этот подход, Ходжкин и Хаксли продемонстрировали, что эти изменения мембранного потенциала являются результатом регулируемого открытия и закрытия Na + и К + каналы в плазматической мембране. Впоследствии стало возможным изучать активность отдельных ионных каналов с помощью методики patch-clamp , разработанной Erwin Neher и Bert Sakmann в 1976 г.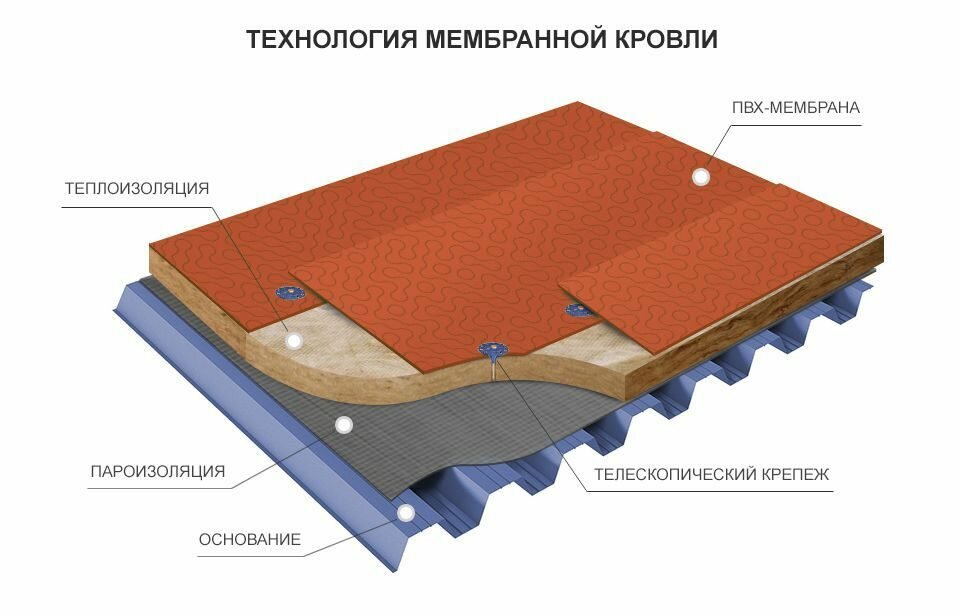 (). В этом методе микропипетка с диаметром наконечника около 1 мкм используется для выделения небольшого участка мембраны, что позволяет анализировать поток ионов через один канал и значительно повышает точность, с которой можно определить активность ионных каналов. учился.
(). В этом методе микропипетка с диаметром наконечника около 1 мкм используется для выделения небольшого участка мембраны, что позволяет анализировать поток ионов через один канал и значительно повышает точность, с которой можно определить активность ионных каналов. учился.
Рисунок 12.19
Метод накладного зажима. На кончике микропипетки выделяют небольшой участок мембраны. Затем внутри пипетки можно применять стимулы, что позволяет измерить поведение захваченного канала. (Адаптировано из E. Neher and B. Sakmann, 1992. (подробнее…)
Поток ионов через мембранные каналы зависит от установления ионных градиентов через плазматическую мембрану. Все клетки, включая нервные и мышечные, содержат ионных насосов (обсуждается в следующем разделе), которые используют энергию, полученную в результате гидролиза АТФ, для активного транспорта ионов через плазматическую мембрану.В результате ионный состав цитоплазмы существенно отличается от внеклеточной жидкости ().Например, Na + активно выкачивается из клеток, а К + закачивается. Таким образом, в аксоне кальмара концентрация Na + примерно в 10 раз выше во внеклеточной жидкости, чем внутри клетки, тогда как концентрация К + примерно в 20 раз выше в цитозоле, чем в окружающей среде.
Таким образом, в аксоне кальмара концентрация Na + примерно в 10 раз выше во внеклеточной жидкости, чем внутри клетки, тогда как концентрация К + примерно в 20 раз выше в цитозоле, чем в окружающей среде.
Таблица 12.1
Концентрации внеклеточных и внутриклеточных ионов.
Поскольку ионы электрически заряжены, их перенос приводит к возникновению электрического градиента на плазматической мембране. В покоящихся аксонах кальмара существует электрический потенциал около 60 мВ через плазматическую мембрану, при этом внутренняя часть клетки отрицательная по отношению к внешней (). Этот электрический потенциал возникает как от ионных насосов, так и от потока ионов через каналы, открытые в плазматической мембране покоящейся клетки. Плазматическая мембрана аксонов покоящегося кальмара содержит открытые K + , поэтому он более проницаем для K + , чем для Na + или других ионов. Следовательно, поток К + вносит наибольший вклад в мембранный потенциал покоя.
Рис. 12.20
Ионные градиенты и мембранный потенциал покоя аксона гигантского кальмара. Показаны только концентрации Na + и K + , поскольку именно эти ионы участвуют в передаче нервных импульсов. Na + выкачивается из клетки, а K + закачивается, (подробнее…)
Как обсуждалось в главе 10, поток ионов через мембрану управляется как концентрацией, так и компонентами напряжения электрохимического градиента. Например, концентрация K + внутри аксона кальмара в 20 раз выше, чем во внеклеточной жидкости, что приводит к выходу K + из клетки. Однако, поскольку K + заряжен положительно, этот выход K + из клетки генерирует электрический потенциал через мембрану, при этом внутренняя часть клетки становится отрицательно заряженной. Этот мембранный потенциал противостоит продолжающемуся потоку K + из клетки, и система приближается к равновесному состоянию, при котором мембранный потенциал уравновешивает градиент концентрации К + .
Количественно связь между концентрацией ионов и мембранным потенциалом определяется уравнением Нернста:
где В – равновесный потенциал в вольтах, R – газовая постоянная, T – абсолютная температура, z – заряд иона, F — постоянная Фарадея, а C o и C i — концентрации иона снаружи и внутри клетки соответственно. Равновесный потенциал существует отдельно для каждого иона, а мембранный потенциал определяется потоком всех ионов, пересекающих плазматическую мембрану. Однако, поскольку покоящиеся аксоны кальмаров более проницаемы для K + , чем для Na + или других ионов (включая Cl – ), мембранный потенциал покоя (-60 мВ) близок к равновесному потенциалу, определяемому внутриклеточным и внеклеточный K + концентрации (-75 мВ).
Когда нервные импульсы ( потенциалы действия ) проходят по аксонам, мембрана деполяризуется (). Мембранный потенциал изменяется от -60 мВ до примерно +30 мВ менее чем за миллисекунду, после чего снова становится отрицательным и возвращается к своему значению покоя. Эти изменения являются результатом быстрого последовательного открытия и закрытия потенциалзависимых каналов Na + и K + . Относительно небольшие начальные изменения мембранного потенциала (от -60 до примерно -40 мВ) приводят к быстрому открытию Na + канала. Это позволяет Na + поступать в клетку под действием как градиента его концентрации, так и мембранного потенциала. Внезапное поступление Na + приводит к сильному изменению мембранного потенциала, который увеличивается почти до +30 мВ, приближаясь к равновесному потенциалу Na + примерно +50 мВ. В это время каналы Na + инактивируются, а потенциалзависимые каналы K + открываются, существенно повышая проницаемость мембраны для K + . Затем K + быстро вытекает из клетки под действием как мембранного потенциала, так и градиента концентрации K + , что приводит к быстрому снижению мембранного потенциала примерно до -75 мВ (равновесный потенциал K + ).
Эти изменения являются результатом быстрого последовательного открытия и закрытия потенциалзависимых каналов Na + и K + . Относительно небольшие начальные изменения мембранного потенциала (от -60 до примерно -40 мВ) приводят к быстрому открытию Na + канала. Это позволяет Na + поступать в клетку под действием как градиента его концентрации, так и мембранного потенциала. Внезапное поступление Na + приводит к сильному изменению мембранного потенциала, который увеличивается почти до +30 мВ, приближаясь к равновесному потенциалу Na + примерно +50 мВ. В это время каналы Na + инактивируются, а потенциалзависимые каналы K + открываются, существенно повышая проницаемость мембраны для K + . Затем K + быстро вытекает из клетки под действием как мембранного потенциала, так и градиента концентрации K + , что приводит к быстрому снижению мембранного потенциала примерно до -75 мВ (равновесный потенциал K + ). Затем потенциалзависимые каналы K + инактивируются, и мембранный потенциал возвращается к уровню покоя -60 мВ, определяемому потоком K + и других ионов через каналы, которые остаются открытыми в нестимулированных клетках.
Затем потенциалзависимые каналы K + инактивируются, и мембранный потенциал возвращается к уровню покоя -60 мВ, определяемому потоком K + и других ионов через каналы, которые остаются открытыми в нестимулированных клетках.
Рис. 12.21
Мембранный потенциал и ионные каналы во время потенциала действия. (A) Изменения мембранного потенциала в одной точке гигантского аксона кальмара после стимула. E Na и E K – равновесные потенциалы Na + и K + соответственно. (B) Мембранный потенциал (подробнее…)
Деполяризация соседних областей плазматической мембраны позволяет потенциалам действия проходить по длине аксонов нервных клеток в виде электрических сигналов, что приводит к быстрой передаче нервных импульсов на большие расстояния. Например, аксоны двигательных нейронов человека могут иметь длину более метра. Прибытие потенциалов действия на окончание большинства нейронов сигнализирует о высвобождении нейротрансмиттеров, таких как ацетилхолин, которые передают сигналы между клетками в синапсах. Нейротрансмиттеры, высвобождаемые из пресинаптических клеток, связываются с рецепторами на мембранах постсинаптических клеток, где они действуют, открывая управляемые лигандом ионные каналы. Одним из наиболее хорошо охарактеризованных таких каналов является ацетилхолиновый рецептор мышечных клеток. Связывание ацетилхолина открывает канал, проницаемый как для Na + и К +. Это обеспечивает быстрый приток Na + , который деполяризует мембрану мышечной клетки и запускает потенциал действия. Затем потенциал действия приводит к открытию потенциалзависимых каналов Ca 2+ , что приводит к увеличению внутриклеточного количества Ca 2+ , что сигнализирует о сокращении (see ).
Нейротрансмиттеры, высвобождаемые из пресинаптических клеток, связываются с рецепторами на мембранах постсинаптических клеток, где они действуют, открывая управляемые лигандом ионные каналы. Одним из наиболее хорошо охарактеризованных таких каналов является ацетилхолиновый рецептор мышечных клеток. Связывание ацетилхолина открывает канал, проницаемый как для Na + и К +. Это обеспечивает быстрый приток Na + , который деполяризует мембрану мышечной клетки и запускает потенциал действия. Затем потенциал действия приводит к открытию потенциалзависимых каналов Ca 2+ , что приводит к увеличению внутриклеточного количества Ca 2+ , что сигнализирует о сокращении (see ).
Рисунок 12.22
Передача сигналов за счет высвобождения нейротрансмиттера в синапсе. Прибытие нервного импульса на окончание нейрона сигнализирует о слиянии синаптических пузырьков с плазматической мембраной, что приводит к высвобождению нейротрансмиттера из пресинаптической клетки в (далее. ..)
..)
Ацетилхолиновый рецептор, первоначально выделенный из электрического органа скатов Torpedo в 1970-х годах, является прототипом лиганд-управляемых каналов. Рецептор состоит из пяти субъединиц, расположенных в виде цилиндра в мембране (). Считается, что в закрытом состоянии поры канала блокируются боковыми цепями гидрофобных аминокислот. Связывание ацетилхолина вызывает конформационные изменения в рецепторе, так что эти гидрофобные боковые цепи смещаются из канала, открывая пору, через которую проходят положительно заряженные ионы, включая Na + и К +. Однако канал остается непроницаемым для отрицательно заряженных ионов, таких как Cl –, поскольку он выстлан отрицательно заряженными аминокислотами.
Рисунок 12.23
Модель рецептора ацетилхолина. Рецептор состоит из пяти субъединиц, расположенных вокруг центральной поры. Связывание ацетилхолина с участком во внеклеточной области рецептора вызывает аллостерические изменения, которые открывают ворота канала. Канал (подробнее…)
Канал (подробнее…)
Большую степень ионной селективности проявляют потенциалзависимые каналы Na + и K + . Каналы Na + более чем в десять раз более проницаемы для Na + , чем для K + , тогда как каналы K + более чем в тысячу раз более проницаемы для K + , чем для Na + . Селективность канала Na + может быть объяснена, по крайней мере, частично, на основе узких пор, которые действуют как размерный фильтр. Ионный радиус Na + (0,95 Å) меньше, чем у K + (1,33 Å), и считается, что пора канала Na + достаточно узкая, чтобы препятствовать прохождению ионов K + или более крупных ( ).
Рисунок 12.24
Ионная селективность каналов Na + . Узкая пора позволяет проходить Na + , связанным с одной молекулой воды, но препятствует прохождению ионов K + или более крупных ионов.
K + каналы также имеют узкие поры, препятствующие прохождению более крупных ионов. Однако, поскольку Na + имеет меньший ионный радиус, это не объясняет избирательной проницаемости этих каналов для K + . Селективность канала K + основана на другом механизме, который был выяснен при определении трехмерной структуры канала K + с помощью рентгеновской кристаллографии в 1998 г. (). Пора канала содержит фильтр с узкой селективностью, выстланный атомами карбонильного кислорода (C=O) из основной цепи полипептида. Когда К 9Ион 0061 + поступает в фильтр селективности, взаимодействия с этими карбонильными атомами кислорода вытесняют молекулы воды, с которыми связан K + , позволяя обезвоженному K + проходить через поры. Напротив, обезвоженный Na + слишком мал, чтобы взаимодействовать с этими карбонильными атомами кислорода в селективном фильтре, который удерживается жестко открытым. Следовательно, Na + остается связанным с молекулами воды в гидратированном комплексе, который слишком велик, чтобы пройти через канал.
Однако, поскольку Na + имеет меньший ионный радиус, это не объясняет избирательной проницаемости этих каналов для K + . Селективность канала K + основана на другом механизме, который был выяснен при определении трехмерной структуры канала K + с помощью рентгеновской кристаллографии в 1998 г. (). Пора канала содержит фильтр с узкой селективностью, выстланный атомами карбонильного кислорода (C=O) из основной цепи полипептида. Когда К 9Ион 0061 + поступает в фильтр селективности, взаимодействия с этими карбонильными атомами кислорода вытесняют молекулы воды, с которыми связан K + , позволяя обезвоженному K + проходить через поры. Напротив, обезвоженный Na + слишком мал, чтобы взаимодействовать с этими карбонильными атомами кислорода в селективном фильтре, который удерживается жестко открытым. Следовательно, Na + остается связанным с молекулами воды в гидратированном комплексе, который слишком велик, чтобы пройти через канал.
Рисунок 12.25
Избирательность каналов K + . Канал К + содержит фильтр узкой селективности, выстланный карбонильными атомами кислорода (С=О). Поры достаточно широки, чтобы обеспечить прохождение обезвоженного K + , из которого все связанные молекулы воды были вытеснены в виде (далее…)
Потенциалзависимых Na + , K + и Ca 2+ канала принадлежат к большому семейству родственных белков (2). Например, последовательность генома C. elegans выявил почти 200 генов, кодирующих ионные каналы, которые, по-видимому, необходимы, чтобы играть различные роли в клеточной передаче сигналов. Каналы K + состоят из четырех идентичных субъединиц, каждая из которых содержит две или шесть трансмембранных α-спиралей. Каналы Na + и Ca 2+ состоят из одной полипептидной цепи, но каждый полипептид содержит четыре повторяющихся домена, соответствующих субъединицам каналов K + . Запирание напряжения опосредовано одной из трансмембранных α-спиралей, которая содержит несколько положительно заряженных аминокислот. Мембранная деполяризация индуцирует перемещение этих положительных зарядов наружу клетки, сдвигая положение этого трансмембранного сегмента и открывая канал. Быстрая инактивация Na + и К + каналы при распространении потенциалов действия затем опосредованы цитоплазматическими участками полипептидной цепи, которые связываются с цитоплазматическим устьем поры канала и препятствуют дальнейшему току ионов ().
Запирание напряжения опосредовано одной из трансмембранных α-спиралей, которая содержит несколько положительно заряженных аминокислот. Мембранная деполяризация индуцирует перемещение этих положительных зарядов наружу клетки, сдвигая положение этого трансмембранного сегмента и открывая канал. Быстрая инактивация Na + и К + каналы при распространении потенциалов действия затем опосредованы цитоплазматическими участками полипептидной цепи, которые связываются с цитоплазматическим устьем поры канала и препятствуют дальнейшему току ионов ().
Рисунок 12.26
Структуры потенциалзависимых катионных каналов. Каналы K + , Na + и Ca 2+ принадлежат к семейству родственных белков. Канал К + образован объединением четырех идентичных субъединиц, одна из которых показана. На 9Канал 0061 + состоит из одного полипептида (подробнее…)
Рисунок 12.27
Инактивация каналов K + и Na + . После потенциалзависимого открытия каналы K + и Na + быстро инактивируются за счет связывания цитоплазматических частей полипептидных цепей с порой. Для канала K + инактивация опосредована шаровой цепью (подробнее…)
После потенциалзависимого открытия каналы K + и Na + быстро инактивируются за счет связывания цитоплазматических частей полипептидных цепей с порой. Для канала K + инактивация опосредована шаровой цепью (подробнее…)
Широкий спектр ионных каналов (включая Ca 2+ и Cl – каналы) реагируют на разные нейротрансмиттеры или открываются и закрываются с разной кинетикой после деполяризации мембраны. Согласованные действия этих многочисленных каналов ответственны за сложность передачи сигналов в нервной системе. Более того, как будет показано в следующей главе, роль ионных каналов не ограничивается электрически возбудимыми нервными и мышечными клетками; они также играют важную роль в передаче сигналов в других типах клеток. Таким образом, регулируемое открытие и закрытие ионных каналов обеспечивает клетки чувствительным и универсальным механизмом реагирования на различные стимулы окружающей среды.
Активный транспорт, управляемый гидролизом АТФ
Чистый поток молекул за счет облегченной диффузии либо через белки-носители, либо через белки-каналы всегда энергетически направлен вниз в направлении, определяемом электрохимическими градиентами через мембрану. Однако во многих случаях клетка должна транспортировать молекулы против градиента их концентрации. При активном транспорте энергия, обеспечиваемая другой связанной реакцией (например, гидролизом АТФ), используется для перемещения молекул в гору в энергетически невыгодном направлении.
Однако во многих случаях клетка должна транспортировать молекулы против градиента их концентрации. При активном транспорте энергия, обеспечиваемая другой связанной реакцией (например, гидролизом АТФ), используется для перемещения молекул в гору в энергетически невыгодном направлении.
Ионные насосы, отвечающие за поддержание градиента ионов через плазматическую мембрану, являются важными примерами активного транспорта, управляемого непосредственно гидролизом АТФ. Как обсуждалось ранее (см.), концентрация Na + примерно в десять раз выше снаружи, чем внутри клеток, тогда как концентрация K + выше внутри, чем снаружи. Эти градиенты ионов поддерживаются насосом Na + -K + (также называемая Na + -K + АТФаза ), которая использует энергию, полученную в результате гидролиза АТФ, для транспорта Na + и K + против их электрохимических градиентов. Этот процесс является результатом конформационных изменений в помпе, вызванных АТФ. Во-первых, ионы Na + связываются с центрами высокого сродства внутри клетки. Это связывание стимулирует гидролиз АТФ и фосфорилирование помпы, вызывая конформационные изменения, которые обнажают Na + – связывает сайты снаружи клетки и снижает их сродство к Na + . Следовательно, связанный Na + высвобождается во внеклеточную жидкость. В то же время на клеточной поверхности экспонируются высокоаффинные сайты связывания К + -. Связывание внеклеточного K + с этими сайтами затем стимулирует гидролиз фосфатной группы, связанной с насосом, что вызывает второе конформационное изменение, открывая сайты связывания K + цитозолю и снижая их аффинность связывания, так что K + высвобождается внутри клетки. Насос имеет три сайта связывания для Na + и два для K + , поэтому каждый цикл переносит через плазматическую мембрану три Na + и два K + за счет одной молекулы АТФ.
Этот процесс является результатом конформационных изменений в помпе, вызванных АТФ. Во-первых, ионы Na + связываются с центрами высокого сродства внутри клетки. Это связывание стимулирует гидролиз АТФ и фосфорилирование помпы, вызывая конформационные изменения, которые обнажают Na + – связывает сайты снаружи клетки и снижает их сродство к Na + . Следовательно, связанный Na + высвобождается во внеклеточную жидкость. В то же время на клеточной поверхности экспонируются высокоаффинные сайты связывания К + -. Связывание внеклеточного K + с этими сайтами затем стимулирует гидролиз фосфатной группы, связанной с насосом, что вызывает второе конформационное изменение, открывая сайты связывания K + цитозолю и снижая их аффинность связывания, так что K + высвобождается внутри клетки. Насос имеет три сайта связывания для Na + и два для K + , поэтому каждый цикл переносит через плазматическую мембрану три Na + и два K + за счет одной молекулы АТФ.
Рисунок 12.28
Модель для работы насоса Na + -K + .
Важность помпы Na + -K + подтверждается тем фактом, что она потребляет почти 25% АТФ, используемого многими животными клетками. Одна из важнейших ролей Na + и К + градиенты, установленные насосом, — это распространение электрических сигналов в нерве и мышце. Как будет сказано ниже, градиент Na + , создаваемый помпой, также используется для обеспечения активного транспорта множества других молекул. Еще одна важная роль насоса Na + -K + в большинстве клеток животных заключается в поддержании осмотического баланса и объема клетки. Цитоплазма содержит высокую концентрацию органических молекул, включая макромолекулы, аминокислоты, сахара и нуклеотиды. В отсутствие противовеса это будет стимулировать поток воды внутрь за счет осмоса, что, если его не остановить, приведет к набуханию и, в конечном итоге, к разрыву клетки. Необходимый противовес обеспечивается градиентами ионов, установленными Na + -К + насос (). В частности, насос устанавливает более высокую концентрацию Na + снаружи, чем внутри клетки. Как уже обсуждалось, поток K + через открытые каналы дополнительно устанавливает электрический потенциал через плазматическую мембрану. Этот мембранный потенциал, в свою очередь, выталкивает Cl – из клетки, поэтому концентрация Cl – (как и Na + ) примерно в десять раз выше во внеклеточной жидкости, чем в цитоплазме. Эти различия в концентрациях ионов уравновешивают высокие концентрации органических молекул внутри клеток, выравнивая осмотическое давление и предотвращая чистый приток воды.
Необходимый противовес обеспечивается градиентами ионов, установленными Na + -К + насос (). В частности, насос устанавливает более высокую концентрацию Na + снаружи, чем внутри клетки. Как уже обсуждалось, поток K + через открытые каналы дополнительно устанавливает электрический потенциал через плазматическую мембрану. Этот мембранный потенциал, в свою очередь, выталкивает Cl – из клетки, поэтому концентрация Cl – (как и Na + ) примерно в десять раз выше во внеклеточной жидкости, чем в цитоплазме. Эти различия в концентрациях ионов уравновешивают высокие концентрации органических молекул внутри клеток, выравнивая осмотическое давление и предотвращая чистый приток воды.
Рис. 12.29
Градиенты ионов через плазматическую мембрану типичной клетки млекопитающих. Концентрации Na + и Cl – выше снаружи, чем внутри клетки, тогда как концентрация К + выше внутри, чем снаружи. Низкие концентрации Na + и Cl – уравновешивают (подробнее…)
Низкие концентрации Na + и Cl – уравновешивают (подробнее…)
Активный транспорт Ca 2+ через плазматическую мембрану управляется насосом Ca 2+ , который структурно родственен до На + -K + насос и аналогичным образом питается от гидролиза АТФ. Насос Ca 2+ транспортирует Ca 2+ из клетки, поэтому внутриклеточные концентрации Ca 2+ чрезвычайно низки: приблизительно 0,1 мкМ, по сравнению с внеклеточными концентрациями около 1 м M . Эта низкая внутриклеточная концентрация Ca 2+ делает клетку чувствительной к небольшому увеличению внутриклеточного уровня Ca 2+ . Такое временное увеличение внутриклеточного Ca 2+ играют важную роль в клеточной передаче сигналов, как уже отмечалось в отношении мышечного сокращения (см. ) и обсуждается далее в следующей главе.
Аналогичные ионные насосы в плазматических мембранах бактерий, дрожжей и растительных клеток ответственны за активный транспорт H + из клетки. Кроме того, H + активно выкачиваются из клеток, выстилающих желудок, что приводит к повышению кислотности желудочного сока. Структурно различные помпы ответственны за активный транспорт H + на лизосомы и эндосомы (см.). Тем не менее, третий тип насоса H + иллюстрируется АТФ-синтазами митохондрий и хлоропластов: в этих случаях насосы можно рассматривать как работающие в обратном направлении, когда движение ионов по электрохимическому градиенту используется для управления синтезом АТФ.
Кроме того, H + активно выкачиваются из клеток, выстилающих желудок, что приводит к повышению кислотности желудочного сока. Структурно различные помпы ответственны за активный транспорт H + на лизосомы и эндосомы (см.). Тем не менее, третий тип насоса H + иллюстрируется АТФ-синтазами митохондрий и хлоропластов: в этих случаях насосы можно рассматривать как работающие в обратном направлении, когда движение ионов по электрохимическому градиенту используется для управления синтезом АТФ.
Самое большое семейство мембранных транспортеров состоит из транспортеров ABC, названных так потому, что они характеризуются высококонсервативными АТФ-связывающими доменами или A TP- b inding c активы (). Как в прокариотических, так и в эукариотических клетках идентифицировано более сотни членов семейства. У бактерий транспортеры ABC используют энергию, полученную в результате гидролиза АТФ, для транспорта широкого спектра молекул, включая ионы, сахара и аминокислоты. В эукариотических клетках был обнаружен первый переносчик ABC как продукт гена (называемого геном множественной лекарственной устойчивости, или mdr ), который делает раковые клетки устойчивыми к различным лекарствам, используемым в химиотерапии. В настоящее время идентифицированы два переносчика MDR. Обычно они экспрессируются в различных клетках, где они удаляют потенциально токсичные чужеродные соединения. Например, экспрессия переносчика MDR в капиллярных эндотелиальных клетках головного мозга, по-видимому, играет важную роль в защите мозга от токсичных химических веществ. К сожалению, транспортеры MDR часто экспрессируются на высоких уровнях в раковых клетках, где они распознают различные лекарства и выкачивают их из клеток, тем самым делая раковые клетки устойчивыми к широкому спектру химиотерапевтических агентов и создавая серьезное препятствие для успешного лечения рака. .
В эукариотических клетках был обнаружен первый переносчик ABC как продукт гена (называемого геном множественной лекарственной устойчивости, или mdr ), который делает раковые клетки устойчивыми к различным лекарствам, используемым в химиотерапии. В настоящее время идентифицированы два переносчика MDR. Обычно они экспрессируются в различных клетках, где они удаляют потенциально токсичные чужеродные соединения. Например, экспрессия переносчика MDR в капиллярных эндотелиальных клетках головного мозга, по-видимому, играет важную роль в защите мозга от токсичных химических веществ. К сожалению, транспортеры MDR часто экспрессируются на высоких уровнях в раковых клетках, где они распознают различные лекарства и выкачивают их из клеток, тем самым делая раковые клетки устойчивыми к широкому спектру химиотерапевтических агентов и создавая серьезное препятствие для успешного лечения рака. .
Рисунок 12.30
Структура транспортера ABC. Базовая структурная единица транспортеров ABC состоит из шести трансмембранных доменов, за которыми следует АТФ-связывающая кассета. В транспортерах плазматической мембраны (таких как показанный транспортер MDR) две из этих единиц являются (подробнее…)
В транспортерах плазматической мембраны (таких как показанный транспортер MDR) две из этих единиц являются (подробнее…)
Еще одним важным с медицинской точки зрения членом семейства транспортеров ABC является ген, ответственный за муковисцидоз. Хотя он является членом семейства ABC, продукт этого гена (называемый регулятором трансмембранной проводимости при муковисцидозе, или CFTR) функционирует как Cl 9.0061- канал в эпителиальных клетках и дефектный транспорт Cl – характерны для заболевания. Канал CFTR Cl – также необычен тем, что для его открытия, по-видимому, требуется как гидролиз АТФ, так и зависимое от цАМФ фосфорилирование. Структурная основа функции CFTR как регулируемого ионного канала еще предстоит выяснить в ходе будущих исследований.
Активный транспорт, управляемый ионными градиентами
Ионные насосы и АВС-переносчики, рассмотренные в предыдущем разделе, используют энергию, полученную непосредственно в результате гидролиза АТФ, для транспортировки молекул против их электрохимических градиентов. Другие молекулы транспортируются против градиента их концентрации с использованием энергии, полученной не от гидролиза АТФ, а от связанного транспорта второй молекулы в энергетически выгодном направлении. На 9Градиент 0061 + , созданный насосом Na + -K + , обеспечивает источник энергии, который часто используется для обеспечения активного транспорта сахаров, аминокислот и ионов в клетках млекопитающих. Аналогичную роль играют градиенты H + , установленные насосами H + клеток бактерий, дрожжей и растений.
Другие молекулы транспортируются против градиента их концентрации с использованием энергии, полученной не от гидролиза АТФ, а от связанного транспорта второй молекулы в энергетически выгодном направлении. На 9Градиент 0061 + , созданный насосом Na + -K + , обеспечивает источник энергии, который часто используется для обеспечения активного транспорта сахаров, аминокислот и ионов в клетках млекопитающих. Аналогичную роль играют градиенты H + , установленные насосами H + клеток бактерий, дрожжей и растений.
Эпителиальные клетки, выстилающие кишечник, представляют собой хороший пример активного транспорта, управляемого градиентом Na + . Эти клетки используют активные транспортные системы в апикальных областях своих плазматических мембран для захвата пищевых сахаров и аминокислот из просвета кишечника. Поглощение глюкозы, например, осуществляется транспортером, который координированно транспортирует два Na + и одна глюкоза в клетку (). Поток Na + по его электрохимическому градиенту обеспечивает энергию, необходимую для поглощения пищевой глюкозы и накопления высоких концентраций внутриклеточной глюкозы. Затем глюкоза высвобождается в подлежащую соединительную ткань (которая содержит кровеносные капилляры) на базолатеральной поверхности кишечного эпителия, где она транспортируется по градиенту концентрации за счет облегченной диффузии (12). Таким образом, поглощение глюкозы из просвета кишечника и ее высвобождение в кровоток является хорошим примером поляризованной функции эпителиальных клеток, которая является результатом специфической локализации активного транспорта и облегченной диффузии переносчиков в апикальный и базолатеральный домены плазматической мембраны. , соответственно.
Поток Na + по его электрохимическому градиенту обеспечивает энергию, необходимую для поглощения пищевой глюкозы и накопления высоких концентраций внутриклеточной глюкозы. Затем глюкоза высвобождается в подлежащую соединительную ткань (которая содержит кровеносные капилляры) на базолатеральной поверхности кишечного эпителия, где она транспортируется по градиенту концентрации за счет облегченной диффузии (12). Таким образом, поглощение глюкозы из просвета кишечника и ее высвобождение в кровоток является хорошим примером поляризованной функции эпителиальных клеток, которая является результатом специфической локализации активного транспорта и облегченной диффузии переносчиков в апикальный и базолатеральный домены плазматической мембраны. , соответственно.
Рис. 12.31
Активный транспорт глюкозы. Активный транспорт, управляемый градиентом Na + , отвечает за поглощение глюкозы из просвета кишечника. Транспортер координированно связывает и транспортирует в клетку одну глюкозу и две Na + . Транспорт Na + (подробнее…)
Транспорт Na + (подробнее…)
Рис. 12.32
Транспорт глюкозы эпителиальными клетками кишечника. Транспортер в апикальном домене плазматической мембраны отвечает за активное поглощение глюкозы (путем котранспорта с Na + ) из просвета кишечника. В результате пищевая глюкоза поглощается и (подробнее…)
Координированное поглощение глюкозы и Na + является примером симпорта, транспорта двух молекул в одном направлении. Напротив, облегченная диффузия глюкозы является примером унипорта, транспорта только одной молекулы. Активный транспорт также может осуществляться антипортом, при котором две молекулы транспортируются в противоположных направлениях (1). Например, Са 2+ экспортируется из клеток не только насосом Ca 2+ , но и антипортером Na + -Ca 2+ , который транспортирует Na + в клетку и Ca 2+ наружу. Другим примером является обменный белок Na + -H + , который участвует в регуляции внутриклеточного pH. Антипортер Na + -H + связывает транспорт Na + в клетку с экспортом H + , тем самым удаляя избыток H + , образующиеся в результате метаболических реакций и препятствующие закислению цитоплазмы.
Антипортер Na + -H + связывает транспорт Na + в клетку с экспортом H + , тем самым удаляя избыток H + , образующиеся в результате метаболических реакций и препятствующие закислению цитоплазмы.
Рисунок 12.33
Примеры антипорта. Ca 2+ и H + экспортируются из клеток антипортерами, которые связывают их экспорт с энергетически выгодным импортом Na + .
Коробка
Молекулярная медицина: кистозный фиброз.
Транспорт малых молекул — клетка
Внутренний состав клетки сохраняется, поскольку плазматическая мембрана избирательно проницаема для малых молекул. Большинство биологических молекул не способны диффундировать через фосфолипидный бислой, поэтому плазматическая мембрана образует барьер, блокирующий свободный обмен молекул между цитоплазмой и внешней средой клетки. Специфические транспортные белки (белки-носители и белковые каналы) опосредуют избирательное прохождение малых молекул через мембрану, позволяя клетке контролировать состав своей цитоплазмы.
Пассивная диффузия
Простейший механизм пересечения молекулами плазматической мембраны — пассивная диффузия. Во время пассивной диффузии молекула просто растворяется в бислое фосфолипидов, диффундирует через него, а затем растворяется в водном растворе на другой стороне мембраны. Никакие мембранные белки не участвуют, и направление транспорта определяется просто относительными концентрациями молекулы внутри и снаружи клетки. Чистый поток молекул всегда направлен вниз по градиенту их концентрации — из отсека с высокой концентрацией в отсек с более низкой концентрацией молекулы.
Таким образом, пассивная диффузия представляет собой неселективный процесс, при котором любая молекула, способная растворяться в двойном слое фосфолипидов, способна пересекать плазматическую мембрану и уравновешиваться между внутренней и внешней частью клетки. Важно отметить, что только небольшие, относительно гидрофобные молекулы способны диффундировать через бислой фосфолипидов со значительной скоростью. Так, газы (такие как O 2 и CO 2 ), гидрофобные молекулы (такие как бензол) и небольшие полярные, но незаряженные молекулы (такие как H 2 O и этанол) способны диффундировать через плазматическую мембрану. Однако другие биологические молекулы не способны растворяться в гидрофобной внутренней части двойного слоя фосфолипидов. Следовательно, более крупные незаряженные полярные молекулы, такие как глюкоза, не могут пересечь плазматическую мембрану путем пассивной диффузии, как и заряженные молекулы любого размера (включая небольшие ионы, такие как H + , Na + , K + и Cl). – ). Прохождение этих молекул через мембрану вместо этого требует активности специфических транспортных и канальных белков, которые, таким образом, контролируют движение большинства биологических молекул в клетку и из нее.
Так, газы (такие как O 2 и CO 2 ), гидрофобные молекулы (такие как бензол) и небольшие полярные, но незаряженные молекулы (такие как H 2 O и этанол) способны диффундировать через плазматическую мембрану. Однако другие биологические молекулы не способны растворяться в гидрофобной внутренней части двойного слоя фосфолипидов. Следовательно, более крупные незаряженные полярные молекулы, такие как глюкоза, не могут пересечь плазматическую мембрану путем пассивной диффузии, как и заряженные молекулы любого размера (включая небольшие ионы, такие как H + , Na + , K + и Cl). – ). Прохождение этих молекул через мембрану вместо этого требует активности специфических транспортных и канальных белков, которые, таким образом, контролируют движение большинства биологических молекул в клетку и из нее.
Рисунок 12.15
Проницаемость фосфолипидных бислоев. Газы, гидрофобные молекулы и небольшие полярные незаряженные молекулы могут диффундировать через бислои фосфолипидов. Более крупные полярные молекулы и заряженные молекулы не могут.
Более крупные полярные молекулы и заряженные молекулы не могут.
Облегченная диффузия и белки-носители
Облегченная диффузия, как и пассивная, включает движение молекул в направлении, определяемом их относительными концентрациями внутри и снаружи клетки. Никакого внешнего источника энергии не предусмотрено, поэтому молекулы движутся через мембрану в направлении, определяемом их градиентами концентрации и, в случае заряженных молекул, электрическим потенциалом на мембране. Однако облегченная диффузия отличается от пассивной диффузии тем, что транспортируемые молекулы не растворяются в двойном слое фосфолипидов. Вместо этого их прохождение опосредуется белками, которые позволяют транспортируемым молекулам пересекать мембрану, не взаимодействуя непосредственно с ее гидрофобной внутренней частью. Таким образом, облегченная диффузия позволяет полярным и заряженным молекулам, таким как углеводы, аминокислоты, нуклеозиды и ионы, пересекать плазматическую мембрану.
Обычно различают два класса белков, которые опосредуют облегченную диффузию: белки-носители и белки-каналы. Белки-переносчики связывают определенные молекулы для транспортировки на одной стороне мембраны. Затем они претерпевают конформационные изменения, которые позволяют молекуле пройти через мембрану и выйти на другую сторону. Напротив, канальные белки (см. следующий раздел) образуют открытые поры через мембрану, обеспечивая свободную диффузию любой молекулы соответствующего размера и заряда.
Белки-переносчики связывают определенные молекулы для транспортировки на одной стороне мембраны. Затем они претерпевают конформационные изменения, которые позволяют молекуле пройти через мембрану и выйти на другую сторону. Напротив, канальные белки (см. следующий раздел) образуют открытые поры через мембрану, обеспечивая свободную диффузию любой молекулы соответствующего размера и заряда.
Белки-носители отвечают за облегченную диффузию сахаров, аминокислот и нуклеозидов через плазматические мембраны большинства клеток. Поглощение глюкозы, которая служит основным источником метаболической энергии, является одной из наиболее важных транспортных функций плазматической мембраны, а транспортер глюкозы является хорошо изученным примером белка-переносчика. Транспортер глюкозы первоначально был идентифицирован как белок массой 55 кД в эритроцитах человека, в которых он составляет примерно 5% от общего белка мембраны. Последующее выделение и анализ последовательности клона кДНК выявили, что переносчик глюкозы имеет 12 α-спиральных трансмембранных сегментов — структура, типичная для многих белков-переносчиков (12). Эти трансмембранные α-спирали содержат преимущественно гидрофобные аминокислоты, но некоторые из них также содержат полярные аминокислотные остатки, которые, как считается, образуют сайт связывания глюкозы внутри белка.
Эти трансмембранные α-спирали содержат преимущественно гидрофобные аминокислоты, но некоторые из них также содержат полярные аминокислотные остатки, которые, как считается, образуют сайт связывания глюкозы внутри белка.
Рисунок 12.16
Структура переносчика глюкозы. Транспортер глюкозы имеет 12 трансмембранных α-спиралей. Полярные аминокислотные остатки, расположенные внутри фосфолипидного бислоя, обозначены темно-фиолетовыми кружками. (Адаптировано из G. I. Bell, C. F. Burant, J. Takeda (подробнее…)
Как и для многих мембранных белков, трехмерная структура транспортера глюкозы неизвестна, поэтому молекулярный механизм транспорта глюкозы остается открытым вопросом. Однако кинетические исследования показывают, что переносчик глюкозы функционирует путем чередования двух конформационных состояний (1). В первой конформации сайт связывания глюкозы обращен к внешней стороне клетки. Связывание глюкозы с этим внешним сайтом вызывает конформационные изменения в транспортер, так что сайт связывания глюкозы теперь обращен внутрь клетки. Затем глюкоза может высвобождаться в цитозоль, после чего транспортер возвращается к своей исходной конформации.
Затем глюкоза может высвобождаться в цитозоль, после чего транспортер возвращается к своей исходной конформации.
Рисунок 12.17
Модель облегченной диффузии глюкозы. Транспортер глюкозы чередуется между двумя конформациями, в которых сайт связывания глюкозы попеременно экспонируется снаружи и внутри клетки. В первой показанной конформации (A) глюкоза (подробнее…)
Большинство клеток, включая эритроциты, подвергаются воздействию внеклеточных концентраций глюкозы, которые выше, чем концентрации внутри клетки, поэтому облегченная диффузия приводит к чистому транспорту глюкозы внутрь . Как только глюкоза поглощается этими клетками, она быстро метаболизируется, поэтому внутриклеточные концентрации глюкозы остаются низкими, и глюкоза продолжает транспортироваться в клетку из внеклеточной жидкости. Однако, поскольку конформационные изменения переносчика глюкозы обратимы, глюкоза может транспортироваться в противоположном направлении, просто обращая шаги в . Такой обратный поток имеет место, например, в клетках печени, в которых синтезируется глюкоза и высвобождается в кровоток.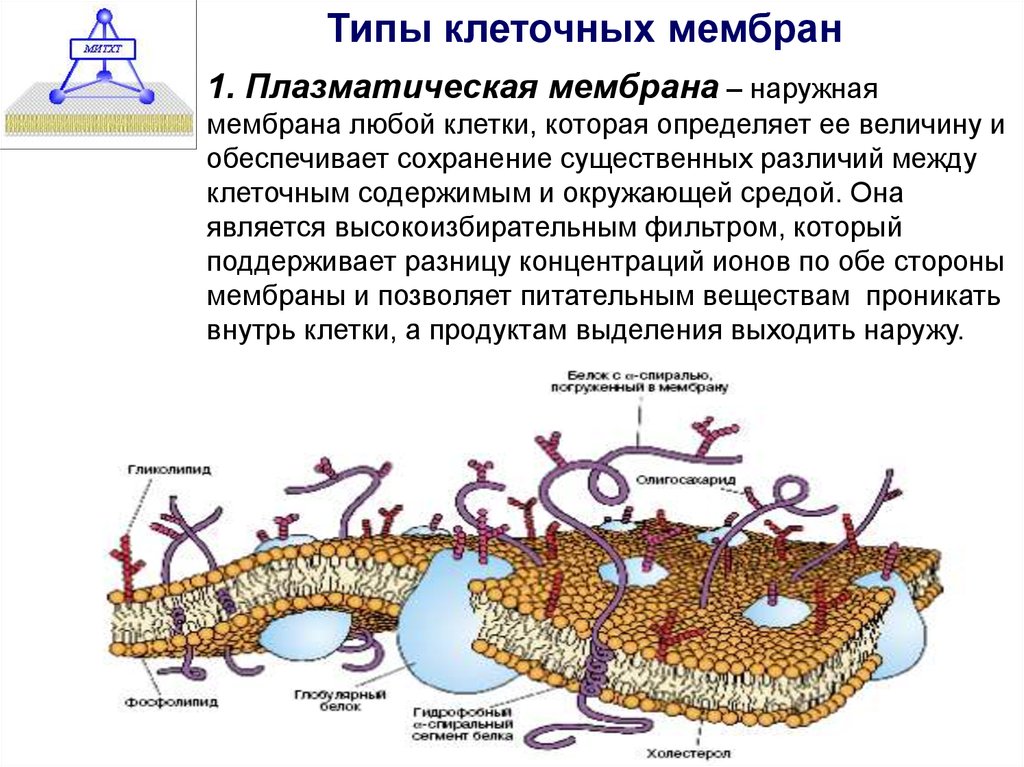
Ионные каналы
В отличие от белков-носителей, канальные белки просто образуют открытые поры в мембране, позволяя небольшим молекулам соответствующего размера и заряда свободно проходить через липидный бислой. Одна группа белков каналов, обсуждавшаяся ранее, — это порины, которые обеспечивают свободное прохождение ионов и небольших полярных молекул через наружные мембраны бактерий (см. ). Канальные белки также обеспечивают прохождение молекул между клетками, соединенными щелевыми контактами, которые обсуждаются далее в этой главе. Плазматические мембраны многих клеток также содержат белки водных каналов (аквапорины), через которые молекулы воды способны пересекать мембрану гораздо быстрее, чем они могут диффундировать через бислой фосфолипидов. Однако наиболее хорошо охарактеризованными белками каналов являются ионные каналы, которые опосредуют прохождение ионов через плазматические мембраны. Хотя ионные каналы присутствуют в мембранах всех клеток, они особенно хорошо изучены в нервах и мышцах, где их регулируемое открытие и закрытие отвечает за передачу электрических сигналов.
Три свойства ионных каналов лежат в основе их функции (). Во-первых, транспортировка по каналам чрезвычайно быстра. Более миллиона ионов в секунду проходят через открытые каналы — скорость потока примерно в тысячу раз превышает скорость транспорта белков-переносчиков. Во-вторых, ионные каналы обладают высокой селективностью, поскольку узкие поры в канале ограничивают прохождение ионов соответствующего размера и заряда. Таким образом, специфические белки каналов обеспечивают прохождение Na + , K + , Ca 2+ и Cl – через мембрану. В-третьих, большинство ионных каналов открыты не постоянно. Вместо этого открытие ионных каналов регулируется «воротами», которые кратковременно открываются в ответ на определенные раздражители. Некоторые каналы (называемые лиганд-зависимыми каналами ) открываются в ответ на связывание нейротрансмиттеров или других сигнальных молекул; другие ( потенциалзависимых канала ) открываются в ответ на изменения электрического потенциала на плазматической мембране.
Рисунок 12.18
Модель ионного канала. В закрытой конформации поток ионов блокируется воротами. Открытие ворот позволяет ионам быстро проходить через канал. Канал содержит узкую пору, которая ограничивает прохождение ионов соответствующего размера (подробнее…)
Фундаментальная роль ионных каналов в передаче электрических импульсов была выяснена в серии элегантных экспериментов, описанных Аланом Ходжкиным и Эндрю Хаксли. в 1952 году. Эти исследователи использовали в качестве модели гигантские нервные клетки кальмара. Аксоны этих гигантских нейронов имеют диаметр около 1 мм, что позволяет вводить электроды и измерять изменения мембранного потенциала, происходящие при передаче нервных импульсов. Используя этот подход, Ходжкин и Хаксли продемонстрировали, что эти изменения мембранного потенциала являются результатом регулируемого открытия и закрытия Na + и К + каналы в плазматической мембране. Впоследствии стало возможным изучать активность отдельных ионных каналов с помощью методики patch-clamp , разработанной Erwin Neher и Bert Sakmann в 1976 г. (). В этом методе микропипетка с диаметром наконечника около 1 мкм используется для выделения небольшого участка мембраны, что позволяет анализировать поток ионов через один канал и значительно повышает точность, с которой можно определить активность ионных каналов. учился.
(). В этом методе микропипетка с диаметром наконечника около 1 мкм используется для выделения небольшого участка мембраны, что позволяет анализировать поток ионов через один канал и значительно повышает точность, с которой можно определить активность ионных каналов. учился.
Рисунок 12.19
Метод накладного зажима. На кончике микропипетки выделяют небольшой участок мембраны. Затем внутри пипетки можно применять стимулы, что позволяет измерить поведение захваченного канала. (Адаптировано из E. Neher and B. Sakmann, 1992. (подробнее…)
Поток ионов через мембранные каналы зависит от установления ионных градиентов через плазматическую мембрану. Все клетки, включая нервные и мышечные, содержат ионных насосов (обсуждается в следующем разделе), которые используют энергию, полученную в результате гидролиза АТФ, для активного транспорта ионов через плазматическую мембрану.В результате ионный состав цитоплазмы существенно отличается от внеклеточной жидкости ().Например, Na + активно выкачивается из клеток, а К + закачивается. Таким образом, в аксоне кальмара концентрация Na + примерно в 10 раз выше во внеклеточной жидкости, чем внутри клетки, тогда как концентрация К + примерно в 20 раз выше в цитозоле, чем в окружающей среде.
Таким образом, в аксоне кальмара концентрация Na + примерно в 10 раз выше во внеклеточной жидкости, чем внутри клетки, тогда как концентрация К + примерно в 20 раз выше в цитозоле, чем в окружающей среде.
Таблица 12.1
Концентрации внеклеточных и внутриклеточных ионов.
Поскольку ионы электрически заряжены, их перенос приводит к возникновению электрического градиента на плазматической мембране. В покоящихся аксонах кальмара существует электрический потенциал около 60 мВ через плазматическую мембрану, при этом внутренняя часть клетки отрицательная по отношению к внешней (). Этот электрический потенциал возникает как от ионных насосов, так и от потока ионов через каналы, открытые в плазматической мембране покоящейся клетки. Плазматическая мембрана аксонов покоящегося кальмара содержит открытые K + , поэтому он более проницаем для K + , чем для Na + или других ионов. Следовательно, поток К + вносит наибольший вклад в мембранный потенциал покоя.
Рис. 12.20
Ионные градиенты и мембранный потенциал покоя аксона гигантского кальмара. Показаны только концентрации Na + и K + , поскольку именно эти ионы участвуют в передаче нервных импульсов. Na + выкачивается из клетки, а K + закачивается, (подробнее…)
Как обсуждалось в главе 10, поток ионов через мембрану управляется как концентрацией, так и компонентами напряжения электрохимического градиента. Например, концентрация K + внутри аксона кальмара в 20 раз выше, чем во внеклеточной жидкости, что приводит к выходу K + из клетки. Однако, поскольку K + заряжен положительно, этот выход K + из клетки генерирует электрический потенциал через мембрану, при этом внутренняя часть клетки становится отрицательно заряженной. Этот мембранный потенциал противостоит продолжающемуся потоку K + из клетки, и система приближается к равновесному состоянию, при котором мембранный потенциал уравновешивает градиент концентрации К + .
Количественно связь между концентрацией ионов и мембранным потенциалом определяется уравнением Нернста:
где В – равновесный потенциал в вольтах, R – газовая постоянная, T – абсолютная температура, z – заряд иона, F — постоянная Фарадея, а C o и C i — концентрации иона снаружи и внутри клетки соответственно. Равновесный потенциал существует отдельно для каждого иона, а мембранный потенциал определяется потоком всех ионов, пересекающих плазматическую мембрану. Однако, поскольку покоящиеся аксоны кальмаров более проницаемы для K + , чем для Na + или других ионов (включая Cl – ), мембранный потенциал покоя (-60 мВ) близок к равновесному потенциалу, определяемому внутриклеточным и внеклеточный K + концентрации (-75 мВ).
Когда нервные импульсы ( потенциалы действия ) проходят по аксонам, мембрана деполяризуется (). Мембранный потенциал изменяется от -60 мВ до примерно +30 мВ менее чем за миллисекунду, после чего снова становится отрицательным и возвращается к своему значению покоя. Эти изменения являются результатом быстрого последовательного открытия и закрытия потенциалзависимых каналов Na + и K + . Относительно небольшие начальные изменения мембранного потенциала (от -60 до примерно -40 мВ) приводят к быстрому открытию Na + канала. Это позволяет Na + поступать в клетку под действием как градиента его концентрации, так и мембранного потенциала. Внезапное поступление Na + приводит к сильному изменению мембранного потенциала, который увеличивается почти до +30 мВ, приближаясь к равновесному потенциалу Na + примерно +50 мВ. В это время каналы Na + инактивируются, а потенциалзависимые каналы K + открываются, существенно повышая проницаемость мембраны для K + . Затем K + быстро вытекает из клетки под действием как мембранного потенциала, так и градиента концентрации K + , что приводит к быстрому снижению мембранного потенциала примерно до -75 мВ (равновесный потенциал K + ).
Эти изменения являются результатом быстрого последовательного открытия и закрытия потенциалзависимых каналов Na + и K + . Относительно небольшие начальные изменения мембранного потенциала (от -60 до примерно -40 мВ) приводят к быстрому открытию Na + канала. Это позволяет Na + поступать в клетку под действием как градиента его концентрации, так и мембранного потенциала. Внезапное поступление Na + приводит к сильному изменению мембранного потенциала, который увеличивается почти до +30 мВ, приближаясь к равновесному потенциалу Na + примерно +50 мВ. В это время каналы Na + инактивируются, а потенциалзависимые каналы K + открываются, существенно повышая проницаемость мембраны для K + . Затем K + быстро вытекает из клетки под действием как мембранного потенциала, так и градиента концентрации K + , что приводит к быстрому снижению мембранного потенциала примерно до -75 мВ (равновесный потенциал K + ). Затем потенциалзависимые каналы K + инактивируются, и мембранный потенциал возвращается к уровню покоя -60 мВ, определяемому потоком K + и других ионов через каналы, которые остаются открытыми в нестимулированных клетках.
Затем потенциалзависимые каналы K + инактивируются, и мембранный потенциал возвращается к уровню покоя -60 мВ, определяемому потоком K + и других ионов через каналы, которые остаются открытыми в нестимулированных клетках.
Рис. 12.21
Мембранный потенциал и ионные каналы во время потенциала действия. (A) Изменения мембранного потенциала в одной точке гигантского аксона кальмара после стимула. E Na и E K – равновесные потенциалы Na + и K + соответственно. (B) Мембранный потенциал (подробнее…)
Деполяризация соседних областей плазматической мембраны позволяет потенциалам действия проходить по длине аксонов нервных клеток в виде электрических сигналов, что приводит к быстрой передаче нервных импульсов на большие расстояния. Например, аксоны двигательных нейронов человека могут иметь длину более метра. Прибытие потенциалов действия на окончание большинства нейронов сигнализирует о высвобождении нейротрансмиттеров, таких как ацетилхолин, которые передают сигналы между клетками в синапсах. Нейротрансмиттеры, высвобождаемые из пресинаптических клеток, связываются с рецепторами на мембранах постсинаптических клеток, где они действуют, открывая управляемые лигандом ионные каналы. Одним из наиболее хорошо охарактеризованных таких каналов является ацетилхолиновый рецептор мышечных клеток. Связывание ацетилхолина открывает канал, проницаемый как для Na + и К +. Это обеспечивает быстрый приток Na + , который деполяризует мембрану мышечной клетки и запускает потенциал действия. Затем потенциал действия приводит к открытию потенциалзависимых каналов Ca 2+ , что приводит к увеличению внутриклеточного количества Ca 2+ , что сигнализирует о сокращении (see ).
Нейротрансмиттеры, высвобождаемые из пресинаптических клеток, связываются с рецепторами на мембранах постсинаптических клеток, где они действуют, открывая управляемые лигандом ионные каналы. Одним из наиболее хорошо охарактеризованных таких каналов является ацетилхолиновый рецептор мышечных клеток. Связывание ацетилхолина открывает канал, проницаемый как для Na + и К +. Это обеспечивает быстрый приток Na + , который деполяризует мембрану мышечной клетки и запускает потенциал действия. Затем потенциал действия приводит к открытию потенциалзависимых каналов Ca 2+ , что приводит к увеличению внутриклеточного количества Ca 2+ , что сигнализирует о сокращении (see ).
Рисунок 12.22
Передача сигналов за счет высвобождения нейротрансмиттера в синапсе. Прибытие нервного импульса на окончание нейрона сигнализирует о слиянии синаптических пузырьков с плазматической мембраной, что приводит к высвобождению нейротрансмиттера из пресинаптической клетки в (далее. ..)
..)
Ацетилхолиновый рецептор, первоначально выделенный из электрического органа скатов Torpedo в 1970-х годах, является прототипом лиганд-управляемых каналов. Рецептор состоит из пяти субъединиц, расположенных в виде цилиндра в мембране (). Считается, что в закрытом состоянии поры канала блокируются боковыми цепями гидрофобных аминокислот. Связывание ацетилхолина вызывает конформационные изменения в рецепторе, так что эти гидрофобные боковые цепи смещаются из канала, открывая пору, через которую проходят положительно заряженные ионы, включая Na + и К +. Однако канал остается непроницаемым для отрицательно заряженных ионов, таких как Cl –, поскольку он выстлан отрицательно заряженными аминокислотами.
Рисунок 12.23
Модель рецептора ацетилхолина. Рецептор состоит из пяти субъединиц, расположенных вокруг центральной поры. Связывание ацетилхолина с участком во внеклеточной области рецептора вызывает аллостерические изменения, которые открывают ворота канала. Канал (подробнее…)
Канал (подробнее…)
Большую степень ионной селективности проявляют потенциалзависимые каналы Na + и K + . Каналы Na + более чем в десять раз более проницаемы для Na + , чем для K + , тогда как каналы K + более чем в тысячу раз более проницаемы для K + , чем для Na + . Селективность канала Na + может быть объяснена, по крайней мере, частично, на основе узких пор, которые действуют как размерный фильтр. Ионный радиус Na + (0,95 Å) меньше, чем у K + (1,33 Å), и считается, что пора канала Na + достаточно узкая, чтобы препятствовать прохождению ионов K + или более крупных ( ).
Рисунок 12.24
Ионная селективность каналов Na + . Узкая пора позволяет проходить Na + , связанным с одной молекулой воды, но препятствует прохождению ионов K + или более крупных ионов.
K + каналы также имеют узкие поры, препятствующие прохождению более крупных ионов. Однако, поскольку Na + имеет меньший ионный радиус, это не объясняет избирательной проницаемости этих каналов для K + . Селективность канала K + основана на другом механизме, который был выяснен при определении трехмерной структуры канала K + с помощью рентгеновской кристаллографии в 1998 г. (). Пора канала содержит фильтр с узкой селективностью, выстланный атомами карбонильного кислорода (C=O) из основной цепи полипептида. Когда К 9Ион 0061 + поступает в фильтр селективности, взаимодействия с этими карбонильными атомами кислорода вытесняют молекулы воды, с которыми связан K + , позволяя обезвоженному K + проходить через поры. Напротив, обезвоженный Na + слишком мал, чтобы взаимодействовать с этими карбонильными атомами кислорода в селективном фильтре, который удерживается жестко открытым. Следовательно, Na + остается связанным с молекулами воды в гидратированном комплексе, который слишком велик, чтобы пройти через канал.
Однако, поскольку Na + имеет меньший ионный радиус, это не объясняет избирательной проницаемости этих каналов для K + . Селективность канала K + основана на другом механизме, который был выяснен при определении трехмерной структуры канала K + с помощью рентгеновской кристаллографии в 1998 г. (). Пора канала содержит фильтр с узкой селективностью, выстланный атомами карбонильного кислорода (C=O) из основной цепи полипептида. Когда К 9Ион 0061 + поступает в фильтр селективности, взаимодействия с этими карбонильными атомами кислорода вытесняют молекулы воды, с которыми связан K + , позволяя обезвоженному K + проходить через поры. Напротив, обезвоженный Na + слишком мал, чтобы взаимодействовать с этими карбонильными атомами кислорода в селективном фильтре, который удерживается жестко открытым. Следовательно, Na + остается связанным с молекулами воды в гидратированном комплексе, который слишком велик, чтобы пройти через канал.
Рисунок 12.25
Избирательность каналов K + . Канал К + содержит фильтр узкой селективности, выстланный карбонильными атомами кислорода (С=О). Поры достаточно широки, чтобы обеспечить прохождение обезвоженного K + , из которого все связанные молекулы воды были вытеснены в виде (далее…)
Потенциалзависимых Na + , K + и Ca 2+ канала принадлежат к большому семейству родственных белков (2). Например, последовательность генома C. elegans выявил почти 200 генов, кодирующих ионные каналы, которые, по-видимому, необходимы, чтобы играть различные роли в клеточной передаче сигналов. Каналы K + состоят из четырех идентичных субъединиц, каждая из которых содержит две или шесть трансмембранных α-спиралей. Каналы Na + и Ca 2+ состоят из одной полипептидной цепи, но каждый полипептид содержит четыре повторяющихся домена, соответствующих субъединицам каналов K + . Запирание напряжения опосредовано одной из трансмембранных α-спиралей, которая содержит несколько положительно заряженных аминокислот. Мембранная деполяризация индуцирует перемещение этих положительных зарядов наружу клетки, сдвигая положение этого трансмембранного сегмента и открывая канал. Быстрая инактивация Na + и К + каналы при распространении потенциалов действия затем опосредованы цитоплазматическими участками полипептидной цепи, которые связываются с цитоплазматическим устьем поры канала и препятствуют дальнейшему току ионов ().
Запирание напряжения опосредовано одной из трансмембранных α-спиралей, которая содержит несколько положительно заряженных аминокислот. Мембранная деполяризация индуцирует перемещение этих положительных зарядов наружу клетки, сдвигая положение этого трансмембранного сегмента и открывая канал. Быстрая инактивация Na + и К + каналы при распространении потенциалов действия затем опосредованы цитоплазматическими участками полипептидной цепи, которые связываются с цитоплазматическим устьем поры канала и препятствуют дальнейшему току ионов ().
Рисунок 12.26
Структуры потенциалзависимых катионных каналов. Каналы K + , Na + и Ca 2+ принадлежат к семейству родственных белков. Канал К + образован объединением четырех идентичных субъединиц, одна из которых показана. На 9Канал 0061 + состоит из одного полипептида (подробнее…)
Рисунок 12.27
Инактивация каналов K + и Na + . После потенциалзависимого открытия каналы K + и Na + быстро инактивируются за счет связывания цитоплазматических частей полипептидных цепей с порой. Для канала K + инактивация опосредована шаровой цепью (подробнее…)
После потенциалзависимого открытия каналы K + и Na + быстро инактивируются за счет связывания цитоплазматических частей полипептидных цепей с порой. Для канала K + инактивация опосредована шаровой цепью (подробнее…)
Широкий спектр ионных каналов (включая Ca 2+ и Cl – каналы) реагируют на разные нейротрансмиттеры или открываются и закрываются с разной кинетикой после деполяризации мембраны. Согласованные действия этих многочисленных каналов ответственны за сложность передачи сигналов в нервной системе. Более того, как будет показано в следующей главе, роль ионных каналов не ограничивается электрически возбудимыми нервными и мышечными клетками; они также играют важную роль в передаче сигналов в других типах клеток. Таким образом, регулируемое открытие и закрытие ионных каналов обеспечивает клетки чувствительным и универсальным механизмом реагирования на различные стимулы окружающей среды.
Активный транспорт, управляемый гидролизом АТФ
Чистый поток молекул за счет облегченной диффузии либо через белки-носители, либо через белки-каналы всегда энергетически направлен вниз в направлении, определяемом электрохимическими градиентами через мембрану. Однако во многих случаях клетка должна транспортировать молекулы против градиента их концентрации. При активном транспорте энергия, обеспечиваемая другой связанной реакцией (например, гидролизом АТФ), используется для перемещения молекул в гору в энергетически невыгодном направлении.
Однако во многих случаях клетка должна транспортировать молекулы против градиента их концентрации. При активном транспорте энергия, обеспечиваемая другой связанной реакцией (например, гидролизом АТФ), используется для перемещения молекул в гору в энергетически невыгодном направлении.
Ионные насосы, отвечающие за поддержание градиента ионов через плазматическую мембрану, являются важными примерами активного транспорта, управляемого непосредственно гидролизом АТФ. Как обсуждалось ранее (см.), концентрация Na + примерно в десять раз выше снаружи, чем внутри клеток, тогда как концентрация K + выше внутри, чем снаружи. Эти градиенты ионов поддерживаются насосом Na + -K + (также называемая Na + -K + АТФаза ), которая использует энергию, полученную в результате гидролиза АТФ, для транспорта Na + и K + против их электрохимических градиентов. Этот процесс является результатом конформационных изменений в помпе, вызванных АТФ. Во-первых, ионы Na + связываются с центрами высокого сродства внутри клетки. Это связывание стимулирует гидролиз АТФ и фосфорилирование помпы, вызывая конформационные изменения, которые обнажают Na + – связывает сайты снаружи клетки и снижает их сродство к Na + . Следовательно, связанный Na + высвобождается во внеклеточную жидкость. В то же время на клеточной поверхности экспонируются высокоаффинные сайты связывания К + -. Связывание внеклеточного K + с этими сайтами затем стимулирует гидролиз фосфатной группы, связанной с насосом, что вызывает второе конформационное изменение, открывая сайты связывания K + цитозолю и снижая их аффинность связывания, так что K + высвобождается внутри клетки. Насос имеет три сайта связывания для Na + и два для K + , поэтому каждый цикл переносит через плазматическую мембрану три Na + и два K + за счет одной молекулы АТФ.
Этот процесс является результатом конформационных изменений в помпе, вызванных АТФ. Во-первых, ионы Na + связываются с центрами высокого сродства внутри клетки. Это связывание стимулирует гидролиз АТФ и фосфорилирование помпы, вызывая конформационные изменения, которые обнажают Na + – связывает сайты снаружи клетки и снижает их сродство к Na + . Следовательно, связанный Na + высвобождается во внеклеточную жидкость. В то же время на клеточной поверхности экспонируются высокоаффинные сайты связывания К + -. Связывание внеклеточного K + с этими сайтами затем стимулирует гидролиз фосфатной группы, связанной с насосом, что вызывает второе конформационное изменение, открывая сайты связывания K + цитозолю и снижая их аффинность связывания, так что K + высвобождается внутри клетки. Насос имеет три сайта связывания для Na + и два для K + , поэтому каждый цикл переносит через плазматическую мембрану три Na + и два K + за счет одной молекулы АТФ.
Рисунок 12.28
Модель для работы насоса Na + -K + .
Важность помпы Na + -K + подтверждается тем фактом, что она потребляет почти 25% АТФ, используемого многими животными клетками. Одна из важнейших ролей Na + и К + градиенты, установленные насосом, — это распространение электрических сигналов в нерве и мышце. Как будет сказано ниже, градиент Na + , создаваемый помпой, также используется для обеспечения активного транспорта множества других молекул. Еще одна важная роль насоса Na + -K + в большинстве клеток животных заключается в поддержании осмотического баланса и объема клетки. Цитоплазма содержит высокую концентрацию органических молекул, включая макромолекулы, аминокислоты, сахара и нуклеотиды. В отсутствие противовеса это будет стимулировать поток воды внутрь за счет осмоса, что, если его не остановить, приведет к набуханию и, в конечном итоге, к разрыву клетки. Необходимый противовес обеспечивается градиентами ионов, установленными Na + -К + насос (). В частности, насос устанавливает более высокую концентрацию Na + снаружи, чем внутри клетки. Как уже обсуждалось, поток K + через открытые каналы дополнительно устанавливает электрический потенциал через плазматическую мембрану. Этот мембранный потенциал, в свою очередь, выталкивает Cl – из клетки, поэтому концентрация Cl – (как и Na + ) примерно в десять раз выше во внеклеточной жидкости, чем в цитоплазме. Эти различия в концентрациях ионов уравновешивают высокие концентрации органических молекул внутри клеток, выравнивая осмотическое давление и предотвращая чистый приток воды.
Необходимый противовес обеспечивается градиентами ионов, установленными Na + -К + насос (). В частности, насос устанавливает более высокую концентрацию Na + снаружи, чем внутри клетки. Как уже обсуждалось, поток K + через открытые каналы дополнительно устанавливает электрический потенциал через плазматическую мембрану. Этот мембранный потенциал, в свою очередь, выталкивает Cl – из клетки, поэтому концентрация Cl – (как и Na + ) примерно в десять раз выше во внеклеточной жидкости, чем в цитоплазме. Эти различия в концентрациях ионов уравновешивают высокие концентрации органических молекул внутри клеток, выравнивая осмотическое давление и предотвращая чистый приток воды.
Рис. 12.29
Градиенты ионов через плазматическую мембрану типичной клетки млекопитающих. Концентрации Na + и Cl – выше снаружи, чем внутри клетки, тогда как концентрация К + выше внутри, чем снаружи. Низкие концентрации Na + и Cl – уравновешивают (подробнее…)
Низкие концентрации Na + и Cl – уравновешивают (подробнее…)
Активный транспорт Ca 2+ через плазматическую мембрану управляется насосом Ca 2+ , который структурно родственен до На + -K + насос и аналогичным образом питается от гидролиза АТФ. Насос Ca 2+ транспортирует Ca 2+ из клетки, поэтому внутриклеточные концентрации Ca 2+ чрезвычайно низки: приблизительно 0,1 мкМ, по сравнению с внеклеточными концентрациями около 1 м M . Эта низкая внутриклеточная концентрация Ca 2+ делает клетку чувствительной к небольшому увеличению внутриклеточного уровня Ca 2+ . Такое временное увеличение внутриклеточного Ca 2+ играют важную роль в клеточной передаче сигналов, как уже отмечалось в отношении мышечного сокращения (см. ) и обсуждается далее в следующей главе.
Аналогичные ионные насосы в плазматических мембранах бактерий, дрожжей и растительных клеток ответственны за активный транспорт H + из клетки. Кроме того, H + активно выкачиваются из клеток, выстилающих желудок, что приводит к повышению кислотности желудочного сока. Структурно различные помпы ответственны за активный транспорт H + на лизосомы и эндосомы (см.). Тем не менее, третий тип насоса H + иллюстрируется АТФ-синтазами митохондрий и хлоропластов: в этих случаях насосы можно рассматривать как работающие в обратном направлении, когда движение ионов по электрохимическому градиенту используется для управления синтезом АТФ.
Кроме того, H + активно выкачиваются из клеток, выстилающих желудок, что приводит к повышению кислотности желудочного сока. Структурно различные помпы ответственны за активный транспорт H + на лизосомы и эндосомы (см.). Тем не менее, третий тип насоса H + иллюстрируется АТФ-синтазами митохондрий и хлоропластов: в этих случаях насосы можно рассматривать как работающие в обратном направлении, когда движение ионов по электрохимическому градиенту используется для управления синтезом АТФ.
Самое большое семейство мембранных транспортеров состоит из транспортеров ABC, названных так потому, что они характеризуются высококонсервативными АТФ-связывающими доменами или A TP- b inding c активы (). Как в прокариотических, так и в эукариотических клетках идентифицировано более сотни членов семейства. У бактерий транспортеры ABC используют энергию, полученную в результате гидролиза АТФ, для транспорта широкого спектра молекул, включая ионы, сахара и аминокислоты. В эукариотических клетках был обнаружен первый переносчик ABC как продукт гена (называемого геном множественной лекарственной устойчивости, или mdr ), который делает раковые клетки устойчивыми к различным лекарствам, используемым в химиотерапии. В настоящее время идентифицированы два переносчика MDR. Обычно они экспрессируются в различных клетках, где они удаляют потенциально токсичные чужеродные соединения. Например, экспрессия переносчика MDR в капиллярных эндотелиальных клетках головного мозга, по-видимому, играет важную роль в защите мозга от токсичных химических веществ. К сожалению, транспортеры MDR часто экспрессируются на высоких уровнях в раковых клетках, где они распознают различные лекарства и выкачивают их из клеток, тем самым делая раковые клетки устойчивыми к широкому спектру химиотерапевтических агентов и создавая серьезное препятствие для успешного лечения рака. .
В эукариотических клетках был обнаружен первый переносчик ABC как продукт гена (называемого геном множественной лекарственной устойчивости, или mdr ), который делает раковые клетки устойчивыми к различным лекарствам, используемым в химиотерапии. В настоящее время идентифицированы два переносчика MDR. Обычно они экспрессируются в различных клетках, где они удаляют потенциально токсичные чужеродные соединения. Например, экспрессия переносчика MDR в капиллярных эндотелиальных клетках головного мозга, по-видимому, играет важную роль в защите мозга от токсичных химических веществ. К сожалению, транспортеры MDR часто экспрессируются на высоких уровнях в раковых клетках, где они распознают различные лекарства и выкачивают их из клеток, тем самым делая раковые клетки устойчивыми к широкому спектру химиотерапевтических агентов и создавая серьезное препятствие для успешного лечения рака. .
Рисунок 12.30
Структура транспортера ABC. Базовая структурная единица транспортеров ABC состоит из шести трансмембранных доменов, за которыми следует АТФ-связывающая кассета. В транспортерах плазматической мембраны (таких как показанный транспортер MDR) две из этих единиц являются (подробнее…)
В транспортерах плазматической мембраны (таких как показанный транспортер MDR) две из этих единиц являются (подробнее…)
Еще одним важным с медицинской точки зрения членом семейства транспортеров ABC является ген, ответственный за муковисцидоз. Хотя он является членом семейства ABC, продукт этого гена (называемый регулятором трансмембранной проводимости при муковисцидозе, или CFTR) функционирует как Cl 9.0061- канал в эпителиальных клетках и дефектный транспорт Cl – характерны для заболевания. Канал CFTR Cl – также необычен тем, что для его открытия, по-видимому, требуется как гидролиз АТФ, так и зависимое от цАМФ фосфорилирование. Структурная основа функции CFTR как регулируемого ионного канала еще предстоит выяснить в ходе будущих исследований.
Активный транспорт, управляемый ионными градиентами
Ионные насосы и АВС-переносчики, рассмотренные в предыдущем разделе, используют энергию, полученную непосредственно в результате гидролиза АТФ, для транспортировки молекул против их электрохимических градиентов. Другие молекулы транспортируются против градиента их концентрации с использованием энергии, полученной не от гидролиза АТФ, а от связанного транспорта второй молекулы в энергетически выгодном направлении. На 9Градиент 0061 + , созданный насосом Na + -K + , обеспечивает источник энергии, который часто используется для обеспечения активного транспорта сахаров, аминокислот и ионов в клетках млекопитающих. Аналогичную роль играют градиенты H + , установленные насосами H + клеток бактерий, дрожжей и растений.
Эпителиальные клетки, выстилающие кишечник, представляют собой хороший пример активного транспорта, управляемого градиентом Na + . Эти клетки используют активные транспортные системы в апикальных областях своих плазматических мембран для захвата пищевых сахаров и аминокислот из просвета кишечника. Поглощение глюкозы, например, осуществляется транспортером, который координированно транспортирует два Na + и одна глюкоза в клетку (). Поток Na + по его электрохимическому градиенту обеспечивает энергию, необходимую для поглощения пищевой глюкозы и накопления высоких концентраций внутриклеточной глюкозы. Затем глюкоза высвобождается в подлежащую соединительную ткань (которая содержит кровеносные капилляры) на базолатеральной поверхности кишечного эпителия, где она транспортируется по градиенту концентрации за счет облегченной диффузии (12). Таким образом, поглощение глюкозы из просвета кишечника и ее высвобождение в кровоток является хорошим примером поляризованной функции эпителиальных клеток, которая является результатом специфической локализации активного транспорта и облегченной диффузии переносчиков в апикальный и базолатеральный домены плазматической мембраны. , соответственно.
Рис. 12.31
Активный транспорт глюкозы. Активный транспорт, управляемый градиентом Na + , отвечает за поглощение глюкозы из просвета кишечника. Транспортер координированно связывает и транспортирует в клетку одну глюкозу и две Na + . Транспорт Na + (подробнее…)
Рис. 12.32
Транспорт глюкозы эпителиальными клетками кишечника. Транспортер в апикальном домене плазматической мембраны отвечает за активное поглощение глюкозы (путем котранспорта с Na + ) из просвета кишечника. В результате пищевая глюкоза поглощается и (подробнее…)
Координированное поглощение глюкозы и Na + является примером симпорта, транспорта двух молекул в одном направлении. Напротив, облегченная диффузия глюкозы является примером унипорта, транспорта только одной молекулы. Активный транспорт также может осуществляться антипортом, при котором две молекулы транспортируются в противоположных направлениях (1). Например, Са 2+ экспортируется из клеток не только насосом Ca 2+ , но и антипортером Na + -Ca 2+ , который транспортирует Na + в клетку и Ca 2+ наружу. Другим примером является обменный белок Na + -H + , который участвует в регуляции внутриклеточного pH. Антипортер Na + -H + связывает транспорт Na + в клетку с экспортом H + , тем самым удаляя избыток H + , образующиеся в результате метаболических реакций и препятствующие закислению цитоплазмы.
Рисунок 12.33
Примеры антипорта. Ca 2+ и H + экспортируются из клеток антипортерами, которые связывают их экспорт с энергетически выгодным импортом Na + .
Коробка
Молекулярная медицина: кистозный фиброз.
Клеточная мембрана: диффузия, осмос и активный транспорт
Думайте об этом как о привратнике, страже или пограничнике. Несмотря на то, что ее толщина всего от 6 до 10 нанометров и она видна только в электронный микроскоп, клеточная мембрана удерживает цитоплазму клетки на месте и позволяет только избранным материалам входить и выходить из клетки по мере необходимости.
Эта полупроницаемость, или селективная проницаемость, является результатом двойного слоя (бислоя) молекул фосфолипидов с вкраплениями белковых молекул . Внешняя поверхность каждого слоя состоит из плотно упакованных гидрофильных (или водолюбивых) полярных головок. Внутри, между двумя слоями, вы найдете гидрофобные (или боящиеся воды) неполярные хвосты состоящие из цепочек жирных кислот .
Молекулы холестерина между молекулами фосфолипидов придают эластичной мембране стабильность и делают ее менее проницаемой для водорастворимых веществ. И цитоплазма, и матрикс, материал, в котором лежат клетки, состоят в основном из воды. Полярные головки электростатически притягивают поляризованные молекулы воды, а неполярные хвосты лежат между слоями, экранированные от воды и образующие сухой средний слой.
Внутренняя часть мембраны состоит из маслянистых молекул жирных кислот, обладающих электростатической симметрией, или неполяризованный. Липидорастворимые молекулы могут проходить через этот слой, но водорастворимые молекулы, такие как аминокислоты, сахара и белки, не могут, вместо этого перемещаясь через мембрану через транспортные каналы, образованные встроенными белками каналов . Поскольку фосфолипиды имеют как полярные, так и неполярные области, их также называют амфипатическими молекулами.
Авторы и права: Иллюстрация Кэтрин Борн, Массачусетс,
.Клеточная мембрана удерживает клетку вместе и изолирует ее как отдельную функциональную единицу протоплазмы. Хотя он может спонтанно восстанавливать небольшие разрывы, серьезное повреждение мембраны приведет к распаду клетки. Мембрана разборчива в том, какие молекулы она пропускает или выпускает. Позволяет передвигаться через барьер на диффузия, осмос, или активный транспорт.
Распространение
Диффузия — это естественное явление с наблюдаемыми эффектами, подобными Броуновскому движению. Молекулы или другие частицы спонтанно распространяются или мигрируют из областей с более высокой концентрацией в области с более низкой концентрацией, пока не наступит равновесие. В состоянии равновесия диффузия продолжается, но чистый поток уравновешивается, за исключением случайных колебаний.
Это происходит потому, что все молекулы обладают кинетической энергией хаотического движения. Они движутся с высокой скоростью, сталкиваются друг с другом, меняют направление и удаляются от областей с большей концентрацией в области с меньшей концентрацией. Скорость диффузии зависит от массы и температуры молекулы; более легкие и теплые молекулы движутся быстрее.
Диффузия — это одна из форм пассивного транспорта , которая не требует затрат клеточной энергии. Молекула может пассивно диффундировать через клеточную мембрану, если она растворима в липидах, незаряжена и очень мала, или если ей может помочь молекула-носитель. Самостоятельная диффузия очень мелких или жирорастворимых частиц называется простой диффузией. Вспомогательный процесс известен как облегченная диффузия.
Клеточная мембрана позволяет неполярным молекулам (тем, которые плохо связываются с водой) течь из области, где они высоко сконцентрированы, в область, где их концентрация меньше. В мембрану встроены молекулы трансмембранного белка, называемые 9.0253 каналы белков , которые проходят от внешнего слоя к внутреннему слою и создают благоприятные для диффузии отверстия для движения молекул.
Осмос
Осмос — это форма пассивного транспорта, аналогичная диффузии, при которой растворитель перемещается через селективно проницаемую или полупроницаемую мембрану из области с более высокой концентрацией в область с более низкой концентрацией. Растворы состоят из двух частей: растворителя и растворенного вещества.
Обычно в ячейке содержится примерно 1 % солевого раствора, другими словами, 1 % соли (растворенного вещества) и 99 процентов воды (растворитель). Вода представляет собой полярную молекулу, которая не проходит через липидный бислой; однако он достаточно мал, чтобы проходить через поры, образованные белковыми молекулами, большинства клеточных мембран.
Осмос возникает, когда молекулярная концентрация воды на двух сторонах мембраны различается. Мембрана пропускает растворитель (воду), но не пропускает растворенное вещество (частицы, растворенные в воде).
На перенос посредством осмоса влияет концентрация растворенного вещества (количество частиц) в воде . Одна молекула или один ион растворенного вещества вытесняет одну молекулу воды. Осмолярность — это термин, используемый для описания концентрации частиц растворенного вещества на литр. Когда вода диффундирует в клетку, внутри клетки создается гидростатическое давление. В конце концов, давление внутри клетки становится равным внешнему осмотическому давлению и уравновешивается им.
Изотонический раствор имеет ту же концентрацию растворенного вещества и растворителя, что и внутри клетки, поэтому клетка, помещенная в изотонический раствор (обычно это 1-процентный физиологический раствор для человека), испытывает одинаковый поток воды в клетку и из нее, поддержание равновесия.
Гипотонический раствор имеет меньший растворенный потенциал и более высокий водный потенциал, чем внутри клетки.
 Примером может служить 100-процентная дистиллированная вода, в которой меньше растворенных веществ, чем внутри клетки. Следовательно, если человеческую клетку поместить в гипотонический раствор, молекулы диффундируют по градиенту концентрации до тех пор, пока клеточная мембрана не лопнет.
Примером может служить 100-процентная дистиллированная вода, в которой меньше растворенных веществ, чем внутри клетки. Следовательно, если человеческую клетку поместить в гипотонический раствор, молекулы диффундируют по градиенту концентрации до тех пор, пока клеточная мембрана не лопнет.Гипертонический раствор имеет более высокий растворенный потенциал и более низкий водный потенциал, чем внутри клетки. Так мембрану клетки человека помещают в 10-процентный солевой раствор (10-процентная соль и 90 процентов воды) позволяет воде вытекать из клетки (от более высокой концентрации внутри к более низкой концентрации снаружи), тем самым уменьшая ее.
Активный транспорт
Активный транспорт происходит через полупроницаемую мембрану против нормального градиента концентрации, перемещаясь из области меньшей концентрации в область большей концентрации и требуя затрат энергии, высвобождаемой из молекулы АТФ.
Трансмембранные белковые молекулы, встроенные гидрофильными головками во внешний слой мембраны, способны обнаруживать и перемещать соединения через мембрану.