Пенополистирол ПСБ-С 25
Самая распространенная и универсальная марка пенопласта. Благодаря своим сбалансированным техническим свойствам, применяется во многих сферах:
- тепло- и звукоизоляция скатных крыш, внутренних перегородок, трубопроводов, трехслойных стен без воздушного зазора, для улучшения покрытия грунта.
- наружное утепление фасадов зданий, лоджий, балконов, стен
- утепление полов в жилых помещениях под армированную стяжку
- при производстве сэндвич-панелей
- при утеплении каркасных домов
Купить пенополистирол ПСБ-С 25 в Новосибирске, Томске и Кемерово вы можете позвонив по телефону (383) 240-98-98
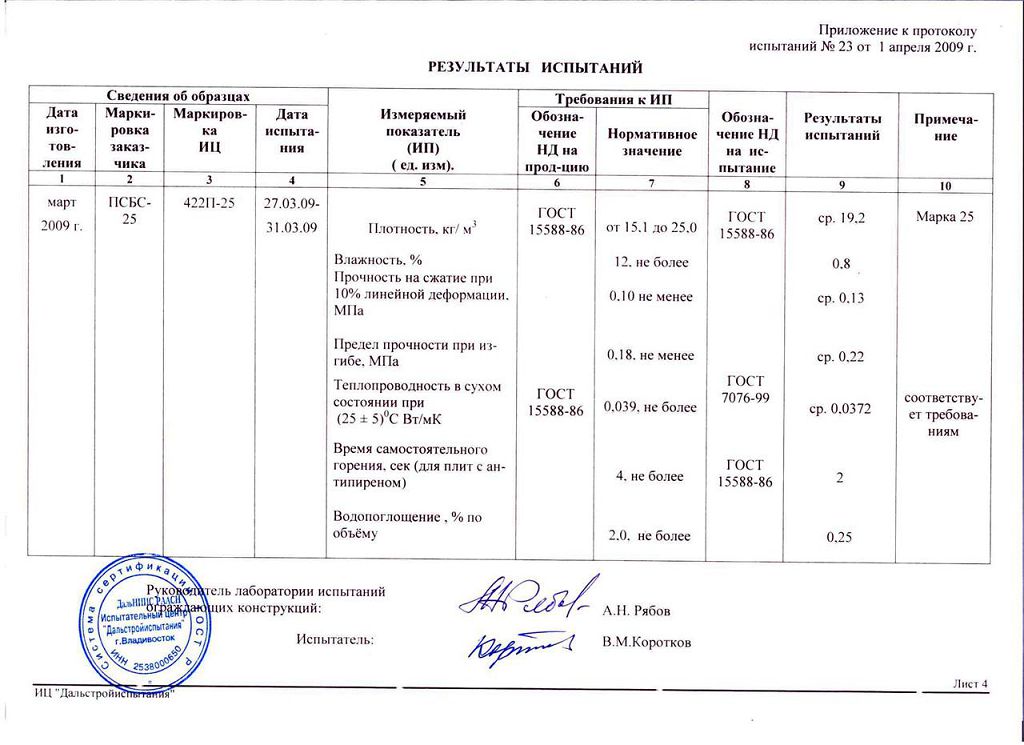
Технические характеристики ПСБ-С 25 (ППС-12,13,14 по ГОСТ 15588-2014)
| Наименованиепоказателя | ЗНАЧЕНИЕ ПОКАЗАТЕЛЯ ДЛЯ ПЛИТ МАРКИ | ||||||||
| ППС10 | ППС12 | ППС13 | ППС14 | ППС16Ф | ППС17 | ППС20 | ППС23 | ППС25 | |
| Плотность, кг/м3, не менее | 10 | 12 | 13 | 14 | 16 | 17 | 20 | 23 | 25 |
| Прочность на сжатие при 10%-ной линейной деформации, кПа, не менее | 40 | 60 | 70 | 80 | 100 | 100 | 120 | 140 | 160 |
| Предел прочности при изгибе, кПа, не менее | 60 | 100 | 120 | 150 | 180 | 160 | 200 | 220 | 250 |
| Предел прочности при растяжении в направлении, перпендикулярном поверхности, кПа, не менее | * | * | * | * | 100 | * | * | * | * |
Теплопроводность плит в сухом состоянии при температуре (10+-1)С (283К), Вт/(м. К), не более К), не более | 0,041 | 0,040 | 0,039 | 0,038 | 0,036 | 0,037 | 0,036 | 0,035 | 0,034 |
| Теплопроводность плит в сухом состоянии при температуре (25+-5)С (298 К), Вт/(м.К), не более | 0,044 | 0,042 | 0,041 | 0,040 | 0,038 | 0,039 | 0,038 | 0,037 | 0,036 |
| Влажность, % по массе, не более | 5,0 | 5,0 | 3,0 | 3,0 | 2,0 | 3,0 | 2,0 | 2,0 | 2,0 |
| Водопоглощение за 24 ч, % по объему, не более | 4,0 | 4,0 | 3,0 | 3,0 | 1,0 | 2,0 | 2,0 | 2,0 | 2,0 |
| время самостоятельного горение, сек, не более | 4 | 4 | 4 | 4 | 1 | 4 | 4 | 4 | 4 |
ПСБ-С-25 пенопласт.
 Реализация продукции ПСБ-С-25, технические характеристики
Реализация продукции ПСБ-С-25, технические характеристикиШирина
Длина
Толщина
Покрытие I
гладкое
Покрытие II
гладкое
Плотность
18.1-18.5 кг/куб.м
Цвет
белый
Прочность
100 кПа
Водопоглощение по объему
до 3%
Горючесть
Г3 (нормальногорючий)
Теплопроводность
0. 038 Вт/мК
038 Вт/мК
Стандарт полный
ГОСТ 15588-1986
Стандарт
Фасовка
1 плита
Упаковка
1 плита
Объем единицы
0.02 куб.м
Объем упаковки
0.02 куб.м
Площадь единицы
1 кв.м
Площадь упаковки
1 кв.![]() м
м
— «ТД КонТРАСТ», Пермь.
Пенополистирол ПСБ-С 25 является пенополистиролом средней плотности и одним из самых распространенных в современном строительстве.
Пенополистирол ПСБ-С 25 — это хороший материал для звуко— и теплоизоляции. Он удобен в монтаже и имеет невысокую стоимость.
Основная сфера применения пенопласта пенополистирола ПСБ-С 25 — это теплоизоляция крыш, стен, перекрытий, а также полов.
ТЕХНИЧЕСКИЕ ХАРАКТЕРИСТИКИ СТИРОПЕН 25 LITE (ПЕНОПЛАСТ ПСБ-С-25)
Марка «Стиропен»
Пенопласт «Стиропен – 25L» | ||
Физико-механические характеристики | Норма по ТУ | факт |
Плотность, кг/м3 | __ | __ |
Прочность на сжатие при 10%-ной линейной деформации, МПа, не менее | 0,08 | 0,08 |
Предел прочности при изгибе, МПа, не менее | 0,16 | 0,16 |
Теплопроводность в сухом состоянии, Вт/м*К, не более | 0,040 | 0,0385 0,0395 |
Время самостоятельного горения, сек. | 4 | 2–3 |
Влажность, %, не более | 12,0 | 0,50–1,10 |
| Водопоглощение за 24 часа, % по объёму, не более | 3,0 | 1,65–2,50 |
 , не более
, не болееТеплоизоляция зданий – одно из важнейших направлений развития современной строительной индустрии.
Применение высококачественных теплоизоляционных материалов Стиропен 25 позволяет снизить массу несущих конструкций, уменьшить потребность в таких строительных материалах как кирпич, бетон, деревянные конструкции. Применение теплоизоляции значительно сокращает энергозатраты при отоплении зданий и позволяет обеспечить максимальный комфорт в жилых помещениях. Кроме того, теплоизоляция высококачественными материалами (пенопласт-пенополистирол) сокращает потребление энергоносителей, то есть различных видов топлива, потребление которых приводит к дополнительным выбросам в атмосферу углекислого газа. Сокращение этих выбросов позволит снизить угрозу глобального потепления.
Сокращение этих выбросов позволит снизить угрозу глобального потепления.
Пенополистирол – экологически чистый, нетоксичный, тепло и звуко изоляционный материал, применяемый в строительстве более 50-ти лет и зарекомендовавший себя как наиболее экономичный, удобный в применении и обладающий низкой теплопроводностью и паропроницаемостью. Поворот к массовому использованию пенополистирола неизбежен, поскольку в обозримом будущем только такие высокоэффективные материалы способны удовлетворить все возрастающие нормативные строительные требования к конструкциям зданий и сооружений Пенополистирол, благодаря своим свойствам, обеспечивает необходимые теплотехнические характеристики строящихся или реконструируемых объектов. Материал на 98% состоит из воздуха – лучшего природного теплоизолятора. Пенополистирол устойчив к воздействию растворов кислот щелочей, спиртов. Инертен по отношению к неорганическим строительным материалам – бетону извести, цементу, песку и др. Разлагается органическими растворителями, смолами, битумным растворами.
Сегодня предприятие выпускает четыре марки пенополистирола строительного блочного самозатухающего в соответствии с ГОСТ 15588-86, отличающиеся по плотности: ПСБ-С-15, ПСБ-С-25, ПСБ-С-25ф, ПСБ-С-35.
Свойства пенополистирола Стиропен 25 Лайт
ППС, благодаря своим свойствам, обеспечивает необходимые теплотехнические характеристики строящихся или реконструированных объектов. Материал на 98% состоит из воздуха – лучшего природного теплоизолятора.
ПСБ-С устойчив к воздействию растворов кислот и щелочей, спиртов. Инертен по отношению к неорганическим строительным материалам – бетону, извести, цементу, песку и др. Разлагается органическими растворами.
Одним из преимуществ пенопласта является способность нести относительно высокую механическую нагрузку при минимальной плотности.
Экологичность
В соответствии с исследованиями института санитарии и гигиены им. Эрисмана пенополистирол не выделяет вредных веществ, а миграция стирола до 85 0С столь незначительна, что не оказывает влияния на здоровье людей. На основании проведенных испытаний нашему предприятию выдано санитарно-эпидемиологическое заключение об экологической безопасности применения ППС в строительстве.
Долговечность
С целью исследования изменений физико-механических и тепло-физических свойств пенополистирола с течением времени проведены ускоренные ресурсные испытания в научно-исследовательском институте строительной физики (г. Москва) по специальной методике с температурными колебаниями от -40 до +40 0С и выдерживанием в воде. Анализ показал, что срок эксплуатации пенополистирольных плит 80 лет.
Пожарные характеристики
Пожарные характеристики являются одним из немаловажных аспектов применения ПСБ-С в строительстве. Сегодня мы имеем сертификат пожарной безопасности продукции по группе горючести Г-2, причем большинство характеристик нашего пенополистирола перекрывают требования группы Г-1. Пожароопасность пенополистирола невелика: теплота, им выделяемая, составляет меньше 2% от всей теплоты пожара. Поэтому в Европе, начиная от Испании и Португалии и заканчивая Норвегией, Финляндией, пенополистирол очень широко применяется в строительстве.
Сегодня мы имеем сертификат пожарной безопасности продукции по группе горючести Г-2, причем большинство характеристик нашего пенополистирола перекрывают требования группы Г-1. Пожароопасность пенополистирола невелика: теплота, им выделяемая, составляет меньше 2% от всей теплоты пожара. Поэтому в Европе, начиная от Испании и Португалии и заканчивая Норвегией, Финляндией, пенополистирол очень широко применяется в строительстве.
В строительстве как утеплитель в ограждающих конструкциях пенополистирол применяется различными способами. Самый очевидный, но не самый лучший – это утепление зданий изнутри. Это сравнительно несложно технологически, но при этом появляется «точка росы» на стыке утеплителя и несущей стены, уменьшается эффективная площадь помещения.
Другие способы — «колодцевая кладка», или «трехслойная кладка». При этом несущая стена рассчитывается только на прочность, а пенополистирол закладывается между несущей конструкцией и облицовочным кирпичом, обеспечивая все теплоизолирующие свойства. Такой способ утепления активно применяется в Удмуртской Республике.
Такой способ утепления активно применяется в Удмуртской Республике.
При третьем, наиболее эффективном с точки зрения теплозащиты и долговечности способе по фасаду здания монтируется система, состоящая из утеплителя, армирующей сетки, клеев, шпаклевок, грунтов, декоративного слоя. Пенополистирол приклеивается к стене, укрепляется дюбелями, а затем наносится армирующая сетка и декоративное покрытие. Так достигается наилучший теплоизолирующий эффект и появляется возможность получить новые архитектурные и дизайнерские решения с помощью пенополистирола.
Область применения
Пенополистирол используется при строительстве холодильных помещений, витрин, морозильных установок, холодильников, емкостей для транспортировки сухого льда и замороженных продуктов, складских помещений и т.д. При применении пенополистирола в холодильной технике учитывается такой показатель, как коэффициент теплопроводности и влагопоглощения, и он по этим свойствам превзошел традиционные теплоизоляционные материалы, ранее используемые в холодильном оборудовании. Пенополистирол имеет закрытую ячеистую структуру, что исключает капиллярное водопоглощение. Такое ценное качество предотвращает его промораживание и разрушение. Он не подвержен гниению. Из этого следует, что срок эксплуатации теплоизоляции из пенополистирола составляет более 80 лет, причем его изоляционные свойства не ухудшаются. Использование пенополистирола при строительстве дорог. Пенополистирол может применяться для распределения нагрузки на дороги и подъезды к мостам в районах, где несущая способность грунта низка. Благодаря небольшому весу, такая конструкция предотвращает опускание дороги.
Пенополистирол имеет закрытую ячеистую структуру, что исключает капиллярное водопоглощение. Такое ценное качество предотвращает его промораживание и разрушение. Он не подвержен гниению. Из этого следует, что срок эксплуатации теплоизоляции из пенополистирола составляет более 80 лет, причем его изоляционные свойства не ухудшаются. Использование пенополистирола при строительстве дорог. Пенополистирол может применяться для распределения нагрузки на дороги и подъезды к мостам в районах, где несущая способность грунта низка. Благодаря небольшому весу, такая конструкция предотвращает опускание дороги.
границ | Ризобактерии, солюбилизирующие фосфат, могут иметь более сильное влияние на свойства корней пшеницы и наземную физиологию, чем солюбилизация ризосферы P
Введение
После азота (N) фосфор (P) является наиболее важным питательным веществом, в котором растения нуждаются в достаточной степени с ранних стадий их развития. Это питательное вещество играет ключевую роль в развитии корней, изменении анатомических свойств корней и густоте корневых волосков, внося значительный вклад в повышение урожайности сельскохозяйственных культур и устойчивости растений к множественным заболеваниям (Kondracka and Rychter, 1997; Ma et al. , 2001). На клеточном уровне P жизненно важен из-за его участия в делении клеток, росте новых тканей и структуре нуклеиновых кислот, которые все регулируют синтез белка, передачу энергии и фотосинтез (Vance et al., 2003). Тем не менее, низкая доступность фосфора в сельскохозяйственных почвах является насущной проблемой, от которой страдают более двух миллиардов гектаров во всем мире (Oberson et al., 2001). Например, сообщалось, что дефицит фосфора вызывает значительное снижение (5-15%) урожайности сельскохозяйственных культур (Shenoy and Kalagudi, 2005) с симптомами дефицита фосфора, характеризующимися красноватыми листьями и некрозом кончиков старых листьев (Luiz et al., 2018). Как правило, доступность фосфора в большинстве почв зависит от множества факторов, в частности от концентрации ионов и pH почвы (Hinsinger et al., 2018). В известковых почвах P часто осаждается вместе с Ca, а в кислых почвах P связывается с Fe и Al (Tariq et al., 2014), в результате чего образуется мало биодоступных форм P для абсорбции корнями, что влияет на рост и продуктивность растений.
, 2001). На клеточном уровне P жизненно важен из-за его участия в делении клеток, росте новых тканей и структуре нуклеиновых кислот, которые все регулируют синтез белка, передачу энергии и фотосинтез (Vance et al., 2003). Тем не менее, низкая доступность фосфора в сельскохозяйственных почвах является насущной проблемой, от которой страдают более двух миллиардов гектаров во всем мире (Oberson et al., 2001). Например, сообщалось, что дефицит фосфора вызывает значительное снижение (5-15%) урожайности сельскохозяйственных культур (Shenoy and Kalagudi, 2005) с симптомами дефицита фосфора, характеризующимися красноватыми листьями и некрозом кончиков старых листьев (Luiz et al., 2018). Как правило, доступность фосфора в большинстве почв зависит от множества факторов, в частности от концентрации ионов и pH почвы (Hinsinger et al., 2018). В известковых почвах P часто осаждается вместе с Ca, а в кислых почвах P связывается с Fe и Al (Tariq et al., 2014), в результате чего образуется мало биодоступных форм P для абсорбции корнями, что влияет на рост и продуктивность растений.
Чтобы преодолеть низкую доступность фосфора в почвах, использование экономичных по фосфору культур наряду с разумным применением различных источников фосфора имеет первостепенное значение для обеспечения урожайности сельскохозяйственных культур.Например, использование видов растений с более высокой способностью поглощать и использовать P в почве с низкой доступностью P было эффективной стратегией, основанной на растениях. Выводы Джаджагло и Рихтера (2008) и Гао и др. (2016) продемонстрировали, что бобовые растения, такие как Medicago sativa , Vicia faba и Phaseolus vulgaris , дают лучшие урожаи в условиях дефицита фосфора и имеют повышенную доступную фракцию фосфора, поглощение фосфора и активность кислых фосфатаз почвы (APase) как следствие более развитая корневая система по сравнению с другими культурами, такими как Sorghum bicolor .Кроме того, диверсификация культур, такая как в случае систем посева бобовых и зерновых, стимулирует поглощение фосфора из-за более высокой подземной биохимической и морфологической функциональной гетерогенности, особенно более быстрого роста корней и более высокой клубеньковости (Hauggaard-Nielsen et al. , 2009; Баргаз и др., 2017). Стимуляция роста растений, доступности и накопления фосфора корнями, вероятно, связана с многочисленными изменениями, вызванными ризосферой, включая закисление ризосферы (Betencourt et al., 2012; Latati et al., 2016), экссудации органических кислот и Р-гидролизующих ферментов (Hakeem et al., 2014), почвенного дыхания (Ibrahim et al., 2013; Latati et al., 2014) и модуляции микробной активности в непосредственной близости от ризосферу (Morgan et al., 2005; Song et al., 2007; Sun et al., 2009). Полезные для сельского хозяйства микроорганизмы, обычно известные как микробы, способствующие росту растений (PGPM), были приняты в качестве мощной микробной стратегии (например, инокулянты, биоудобрения, биостимуляторы, биостимуляторы), которые могут стимулировать рост растений с помощью прямых и / или косвенных механизмов (Mishra and Sundari, 2013; Гупта и др., 2015; Мишра и др., 2017). Прямые эффекты, приписываемые ризобактериям, способствующим росту растений (PGPR), зависят от нескольких физиологических и биохимических путей, которые улучшают питание растений и охватывают большинство механизмов, связанных, среди прочего, с солюбилизацией и поглощением питательных веществ (например, P, K, Zn и т.
, 2009; Баргаз и др., 2017). Стимуляция роста растений, доступности и накопления фосфора корнями, вероятно, связана с многочисленными изменениями, вызванными ризосферой, включая закисление ризосферы (Betencourt et al., 2012; Latati et al., 2016), экссудации органических кислот и Р-гидролизующих ферментов (Hakeem et al., 2014), почвенного дыхания (Ibrahim et al., 2013; Latati et al., 2014) и модуляции микробной активности в непосредственной близости от ризосферу (Morgan et al., 2005; Song et al., 2007; Sun et al., 2009). Полезные для сельского хозяйства микроорганизмы, обычно известные как микробы, способствующие росту растений (PGPM), были приняты в качестве мощной микробной стратегии (например, инокулянты, биоудобрения, биостимуляторы, биостимуляторы), которые могут стимулировать рост растений с помощью прямых и / или косвенных механизмов (Mishra and Sundari, 2013; Гупта и др., 2015; Мишра и др., 2017). Прямые эффекты, приписываемые ризобактериям, способствующим росту растений (PGPR), зависят от нескольких физиологических и биохимических путей, которые улучшают питание растений и охватывают большинство механизмов, связанных, среди прочего, с солюбилизацией и поглощением питательных веществ (например, P, K, Zn и т. Д.). биологический N 2 фиксация и продукция молекул фитогормона и сидерофоров (Fankem et al., 2006; Panhwar et al., 2011). Кроме того, PGPR может косвенно стимулировать рост растений, модулируя местные и системные защитные механизмы растений или производя вторичные метаболиты (аллелохимические вещества), которые действуют как сигналы, индуцирующие иммунитет растений против атак фитопатогенов (Kumar et al., 2018).
Д.). биологический N 2 фиксация и продукция молекул фитогормона и сидерофоров (Fankem et al., 2006; Panhwar et al., 2011). Кроме того, PGPR может косвенно стимулировать рост растений, модулируя местные и системные защитные механизмы растений или производя вторичные метаболиты (аллелохимические вещества), которые действуют как сигналы, индуцирующие иммунитет растений против атак фитопатогенов (Kumar et al., 2018).
Множественные положительные эффекты почвенных микроорганизмов были широко определены как ключевые факторы, способствующие лучшему росту растений и увеличению доступности фосфора в почве (Kumar, 2016; Pérez et al., 2016; Bargaz et al., 2018). PGPR, демонстрирующий высокий уровень PSC, был описан как полезный для роста и урожайности растений, когда он связан с корнями и даже в других частях растения, таких как листья (Fahad et al., 2015; Jambhulkar et al., 2016; Tang et al., 2018). Например, применение эффективных фосфатсолюбилизирующих бактерий (PSB), таких как Bacillus megaterium , повысило доступность фосфора в почве почти на 30% (Alzoubi and Gaibore, 2012).Аналогичным образом, другие виды бактерий, принадлежащих к нескольким родам, таким как Pseudomonas (Sharma et al., 2013), Azotobacter (Kumar and Singh, 2001), Xanthomonas (De Freitas et al., 1997), Rhodococcus , Arthrobacter , Serratia , Chryseobacterium , Gordonia , Phyllobacterium и Delftia sp. (Wani et al., 2005; Chen et al., 2006), как известно, демонстрируют высокий PSC. Помимо однократного использования PSB в качестве биоинокулянтов, двойное использование PSB и минеральных удобрений на основе фосфора, включая малорастворимые формы фосфора, также предоставило доказательства прибыльной интегрированной системы питания растений, которая может привести к успешному производству «микробов и фосфатов». ”Альянс (Аднан и др., 2017; Баргаз и др., 2018; Тахир и др., 2018). Исследования Panhwar et al. (2011) и Bakhshandeh et al. (2015) измерили более высокий урожай риса и подсолнечника в ответ на совместное применение различных минералов P, таких как тройной суперфосфат (TSP), и инокуляции различными PSB ( Bacillus , Rahnella aquatillis , Enterobacter sp., ). Pseudomonas fluorescens и Pseudomonas putida ). Такое положительное двойное использование обоих ресурсов было подтверждено как на физиологических стадиях развития растений, так и на стадиях урожайности зерновых, состоящих из множества функциональных признаков, включая фотосинтетические пигменты -Chl a, Chl b, Chls и Car-, параметры роста, высоту растения, количество метелок холма, высота стеблей, масса зерна, биологический урожай, выход масла из семян, концентрация питательных веществ в семенах и масле.
Комбинированное использование PSB и каменного фосфата (RP), которые считаются природными ресурсами, было успешным благодаря ряду прикладных исследований, которые продемонстрировали повышенную агрономическую эффективность RP (Gomes et al., 2014; Abbasi et al., 2015 ; Giro et al., 2016; Bargaz et al., 2018). Использование микробных функциональных характеристик, связанных с солюбилизацией фосфора, имеет первостепенное значение для предложения микробных стратегий, позволяющих повысить эффективность использования РП, требуемую во многих сельскохозяйственных почвах с высоким удержанием фосфора (Kumar, 2016).Многие экспериментальные исследования предоставили доказательства того, что при объединении штаммов PSB и RP может возникнуть синергизм, что может привести к созданию экономичного биоудобрения на основе фосфора для непосредственного применения в почвах с высоким удержанием фосфора. Например, двойное применение RP и PSB (например, Klebsiella , Azotobacter , Azosporillum и Rhizobium ) значительно улучшило питание фосфора как для зерновых, так и для бобовых культур (Del Pilar López-Ortega et al., 2013; Kaur и Reddy, 2015; Midekssa et al., 2016; Аднан и др., 2017; Manzoor et al., 2017; Дитта и др., 2018). Некоторые параметры наземных и подземных растений используются для количественной оценки таких положительных эффектов; однако эффекты PSB могут дополнять питательные свойства RP, солюбилизация которых должна происходить, не только из-за самих PSB, но также из-за того, в какой степени они могут сильно модулировать как функциональные признаки, так и активность корней. В соответствии с текущими знаниями, тесты in vitro и эксперименты по инокуляции растений были в основном приняты для принятия решений об эффективных бактериальных изолятах PSB, которые будут использоваться в качестве эффективных биоинокулянтов.Тем не менее, поведенческие свойства PSB во временном масштабе на стадиях роста растений необходимо механически распознать и своевременно контролировать либо для одного штамма, либо для консорциума. Это поможет понять, насколько тесна взаимосвязь между интересующим PSB и системой укоренения при умеренно P-формах и всегда ли она остается тесной на разных стадиях роста растений, учитывая, что высокоэффективные PSB in vitro , по-видимому, являются наиболее эффективными . in planta .Другой важный аспект успешного взаимодействия PSB-корень лучше всего подходит с точки зрения стимуляции укоренения в дополнение к солюбилизации ризосферы P, на которой в большинстве исследований было сосредоточено всего несколько исследований (Bakhshandeh et al., 2015; Sarsan, 2016; Rezakhani et al. al., 2019) описали положительное влияние на определенные функциональные особенности корня. Это соответствует цели данного исследования по оценке воздействия пяти ризобактерий, солюбилизирующих P, с различными PSC «низкий, средний и высокий» на морфологические признаки корня твердой пшеницы и связанную с ним солюбилизацию P ризосферы, чтобы пролить свет на то, насколько сильно связывает ли инокуляция параметры ризосферы с морфологическими и физиологическими признаками наземных растений при РП удобрении.
Материалы и методы
Микробные эксперименты
Отбор образцов растений и выделение ризобактерий
В этом исследовании пять изолятов PSB ( Pseudomonas spp.) С контрастирующими PSC использовались в качестве инокулянта для изучения физиологических реакций над и под землей в Пшеница, удобренная RP. Они были среди 42 изолятов PSB, которые были захвачены из ризосферных почв нескольких культур (пшеница, ячмень, кукуруза, овес, фасоль, горох и т. Д.) Из двух основных сельскохозяйственных районов (Хауз и Эрхамна) и из ризосферных почв естественно выращенные растения в районе добычи P в Benguerir в Марокко.Для выделения PSB использовали фосфатную питательную среду Национального института ботанических исследований (NBRIP) -агар с трикальцийфосфатом (TCP, Ca 3 (PO 4 ) 2 , 5 г / л) или RP (5 г / л). содержащий P 2 O 5 : 30,65%, CaO: 48,51%, MgO: 0,63%, K 2 O: 0,09%, Fe 2 O 3 : 0,25%) в качестве единственного источника P добавлено (на литр) глюкозы: 10 г; MgCl 2 ∙ 6H 2 O: 5 г; MgSO 4 ∙ 7H 2 O: 0.25 г; KCl: 0,2 г и (NH 4 ) 2 SO 4 : 0,1 г). Бактериальные изоляты с прозрачными ореолами солюбилизации P хранили как PSB до количественного анализа скорости солюбилизации P в жидкой среде NBRIP с добавлением TCP или RP после 7 дней инкубации при 28 ° C. В дополнение к признаку солюбилизации P изоляты также подвергали скринингу на другие признаки PGP, такие как подкисление среды, фиксация азота, продукция индолуксусной кислоты (IAA), продукция аммония, продукция цианида водорода (HCN) и устойчивость к солености.На основе их PSC все изолированные PSB (включая PSB, испытанные в этом исследовании) были разделены на три группы (низкий, средний и высокий PSC).
Выделение ДНК и молекулярная идентификация
Перед выделением ДНК изоляты PSB культивировали при осторожном перемешивании в 10 мл бульона Лурия-Бертани в течение 24 часов при 28 ° C. Один миллилитр бактериальной культуры помещали в микропробирки и осаждали центрифугированием в течение 2 мин. Для выделения геномной бактериальной ДНК использовали набор GenElute ™ Bacterial Genomic DNA, следуя инструкциям производителя.Бактериальная ДНК из пяти изолятов (от PSB 1 до PSB 5 ) была визуализирована с помощью электрофореза в агарозном геле (0,8%) и количественно определена спектрофотометрически с использованием NanoDrop TM ND-1000 V3.7.0 (Thermo Fisher Scienti c Inc., Уилмингтон, США). перед ПЦР-амплификацией рДНК 16S. Таксономическую идентификацию изолятов проводили путем секвенирования гена 16S рРНК с использованием следующих праймеров: 27F (5’-AGAGTTTGATCCTGGCTCAG-3 ’) и 1492R (5’-GGTTACCTTGTTACGACTT-3’). BLAST-анализ пяти изолятов PSB (от PSB 1 до PSB 5 ) относится к Pseudomonas plecoglossicida , Pseudomonas reinekei , Pseudomonas koreensis , Pseudomonas japonica и freudomonas japonica и соответственно.Последовательности гена 16S рРНК депонированы в GenBank под номерами доступа MT362706 – MT362710.
Определение скорости солюбилизации P
изолята PSB (от PSB 1 до PSB 5 ) были протестированы на их способность солюбилизировать TCP путем определения индекса солюбилизации P (PSI) в агаризованной среде NBRIP после 7 дней инкубации при 28 ° C. ° C. PSI рассчитывали как сумму диаметра колонии и очищенной зоны, деленную на диаметр колонии (Iqbal et al., 2016). Количественную оценку солюбилизации TCP или RP каждым бактериальным изолятом проводили в жидкой среде NBRIP, в которой также отслеживали изменения pH.Среду NBRIP инокулировали 0,1 мл жидкой бактериальной культуры (10 8 КОЕ мл -1 ), инкубировали при 180 об / мин в течение семи дней при 28 ° C, и супернатанты каждой суспензии PSB получали центрифугированием (3000 г за 10 мин). Доступную фракцию P оценивали спектрофотометрически с использованием метода молибденового синего в сравнении со стандартами, нанесенными на спектрофотометр при 880 нм. Оптическую плотность образцов измеряли с помощью стандартной кривой с использованием той же длины волны и переводили в концентрации P, выраженные как мкг ∙ мл -1 (Fernández et al., 2007).
Определение признаков, способствующих росту бактериальных растений
Свободный N 2 Фиксация была подтверждена в среде Эшби, не содержащей азота, состоящей из (на 1 л): агара (15 г), маннита (15 г), K 2 HPO 4 (0,4 г), CaCl 2 • 2H 2 O (0,1 г), NaCl (0,2 г), MgCl 2 (0,1 г ), FeSO 4 • 7H 2 O ( 3,0 мг), NaMoO • 2H 2 O (3,0 мг). После 7 дней инкубации при 28 ° C изоляты PSB, полученные в среде Эшби, считались изолятами со свободным N-фиксатором, и их способность продуцировать аммоний была проверена с использованием реагента Несслера согласно Geetha et al.(2014).
Сначала был проведен качественный анализ продукции ИУК (розовый цвет указывает на продукцию ИУК) в жидкой среде NBRIP с добавлением триптофана с использованием метода Сальковского (Biswas et al., 2018). Во-вторых, затем были отобраны изоляты, продуцирующие ИУК, для оценки продукции ИУК с использованием бактериальных культур, которые выращивали в 50 мл среды и осторожно встряхивали в течение пяти дней при 28 ° C. Два миллилитра реактива Сальковского (смесь 0,5 М хлорида железа (FeCl 3 ) и 35% хлорной кислоты (HClO 4 )) добавляли к 1 мл супернатанта культуры и смесь инкубировали в темноте при комнатной температуре в течение 30 минут. мин.Появление розового цвета, указывающее на продукцию ИУК, которое количественно (оценка) определялось спектрофотометрически при 535 нм с использованием кривой концентрации ИУК, построенной с 0, 10, 20, 50 и 100 мкг ∙ мл -1 синтетической ИУК (Barra et al. , 2016).
Продукция сидерофоров изолятами PSB была выявлена на голубой агаризованной среде CAS (хромазурол S) в соответствии с Pérez-Miranda et al. (2007). После инкубации при 28 ± 2 ° C в течение 5 дней изменение цвета агара CAS с синего на оранжевый вокруг колоний PSB является показателем продукции сидерофоров.Производство цианистого водорода изолятами PSB проводили в среде триптон-соевого агара с добавлением 0,44% глицина (Geetha et al., 2014). После двух дней инкубации при 28 ° C продукция HCN визуально определялась изменением цвета с желтого на красновато-коричневый. Толерантность к засолению была проверена путем выращивания изолятов PSB на среде Лурия-Бертани, снабженной растущими концентрациями NaCl (например, 2, 5 и 8%), инкубированных в течение трех дней при 28 ° C, и устойчивость к засолению определялась простой визуализацией роста бактерий на Luria. –Агаровая среда Бертани (Sarkar et al., 2018).
Эксперимент по инокуляции растений
Инокуляция пшеницы и условий роста растений
Влияние инокуляции изолятами PSB на Радикулы проростков
Пять бактериальных изолятов, демонстрирующих высокий (PSB 3 и PSB 5 ), умеренный (PSB 2 и PSB 4 ) и низкий (PSB 1 ) PSC и множество других PGP-признаков. Их способность улучшать рост проростков пшеницы также определялась у 7-дневных радикальных проростков.Поверхность семян пшеницы стерилизовали гипохлоритом натрия (6 °, 1 мин) и этанолом (96%, 1 мин), а затем тщательно промывали стерильной дистиллированной водой. Инокулят для каждого изолята PSB готовили в жидкой среде Луриа-Бертани при 28 ° C в течение 48 ч (10 8 КОЕ мл -1 ), центрифугировали и осадок клеточных бактерий использовали для посева семян, который вносили путем замачивания семян в 20 мл посевного материала в течение 1 ч при легком встряхивании. Инокулированные семена проращивали в стерилизованной бумаге для проращивания, в которой семена были равномерно распределены, увлажнялись 2 мл стерилизованной воды, смешанной с RP, и скатывались в вертикальном положении бумаги.Семена инкубировали для прорастания в камере для выращивания (фитотрон) в контролируемых условиях (28 ° C, влажность 70%, световой период 16/8 ч и интенсивность освещения 240 мкмоль м -2 с -1 ). Радикулы семидневных проростков измеряли с помощью корневого сканера WinRhizo (Regent Instruments Inc., Квебек, Канада).
Влияние инокуляции изолятами PSB на 15- и 42-дневные растения пшеницы
Семена пшеницы стерилизовали поверхность и инокулировали, как описано выше для параметров прорастания проростков.Вкратце, эксперимент проводился в пластиковых горшках (глубиной 20 см и диаметром 15 см), которые были предварительно стерилизованы (гипохлорит натрия 6 °) и заполнены стерилизованной смесью песка, почвы и торфа (2: 0,5: 0,5). Были протестированы пять обработок с бактериальным посевом (от PSB 1 до PSB 5 ) по сравнению с двумя контрольными обработками. Контрольные обработки соответствуют 1) неинокулированным растениям пшеницы, полученным с каменным P (157 кг га -1 ) и 2) неинокулированным растениям пшеницы, полученным с TSP (85 кг га -1 ) и легкодоступной P-форме ( оценено на основе потребности пшеницы в фосфоре согласно Kaur and Reddy, 2015).Инокулированные обработки (однократная инокуляция изолятами PSB 1 – PSB 5 ) содержали такое же количество либо RP, либо TSP, которые были стерильно смешаны с субстратом для роста растений перед посевом. Эксперимент проводился в контролируемых условиях (28 ° C, влажность 70%, фотопериод 16/8 ч и интенсивность освещения 240 мкмоль м -2 с -1 ) в полной рандомизированной схеме из четырех повторов на обработку с каждая копия состоит из горшка с восемью растениями пшеницы.Орошение производили один раз в неделю раствором Хогланда без фтора и дважды поливали стерильной дистиллированной водой для поддержания адекватной полевой емкости почвы. Через шесть недель после посева были измерены два неразрушающих анализа (например, флуоресценция хлорофилла и устьичная проводимость) (Zeng et al., 2013) перед сбором растений и ризосферных почв для дополнительных наземных и подземных анализов.
Определение параметров растений и ризосферы
Измерение морфологических признаков корней и биомассы растений
Через 15 и 42 дня после прорастания растения собирали и разделяли на побеги и корни.Субстрат для роста ризосферы получали путем осторожного отделения корней от слабо приставшей почвы, которую затем просеивали (2 мм) перед измерениями концентрации фосфора Олсена. Морфологические признаки корней измеряли с использованием программного обеспечения для автоматического анализа изображений WinRhizo (Regent Instruments Inc., Квебек, Канада). Каждый образец корня был равномерно распределен в слое воды на прозрачном лотке из оргстекла и отображен с разрешением 300 dpi с помощью системы сканирования Epson Expression 836 L.Изображения корней анализировали на общую длину корня (RL), площадь поверхности корня (RSA), средний диаметр корня (RD) и объем корня (RV). Впоследствии был определен сухой вес побегов (SDW) и корней (RDW) перед их измельчением в мелкий порошок для анализа концентраций P и N.
Определение доступного фосфора в ризосфере и питательных веществ (P и N) Получение
Доступное содержание фосфора в почве ризосферы было измерено в соответствии с Fernández et al. (2007). Содержание общего фосфора в побегах и корнях определяли в тонко измельченных высушенных образцах (0.5 г), которые сжигали при 600 ° C в течение 6 ч с последующей солюбилизацией золы в соляной кислоте (10 н.). Полученные фильтраты (1 мл) добавляли к 5 мл реакционной смеси, состоящей из молибдата аммония (2,5%) и сульфата гидразина (0,15%), и измеряли оптическую плотность при 820 нм (Majeed et al., 2015). Корни и побеги (100 мг сырой массы (FW)) измельчали экстракционной смесью, состоящей из 500 мкл 0,1 М буфера ацетата натрия (pH 5,6), содержащего 1 мМ дитиотреитол. Гомогенаты центрифугировали при 13000 g при 4 ° C в течение 30 минут и аликвоты по 50 мкл супернатанта использовали для количественного определения неорганического P (Pi) (Bargaz et al., 2012; Баргаз и др., 2017).
Содержание Pi в побегах и корнях измеряли методом аскорбиновой кислоты, как описано Zheng et al. (2009). Части мелко измельченных побегов (0,5 г) также использовались для анализа общего азота с использованием метода Кьельдаля (Magomya et al., 2014). Эффективность получения фосфора корнями (RPAE), которая отражает способность корней поглощать фосфат из почвы, рассчитывалась как отношение содержания фосфора в растении к сухой массе корня (Pan et al., 2008).
Содержание белка и хлорофилла в побегах пшеницы
Образцы 100 мг сырой массы измельчали в 4 мл трис-HCl буфера (0.1 M pH 7,5) и центрифугировали при 15000 g в течение 20 мин. Содержание белка определяли методом Брэдфорда. Концентрацию белка определяли на основе стандартной кривой бычьего сывороточного альбумина. Общая концентрация хлорофилла измерялась согласно Pérez-Patricio et al. (2018). Аликвоту 100 мг свежей ткани листа растирали в 5 мл ацетона (80%, об. / Об.). Общий хлорофилл определяли по следующей формуле:
Chlt = 8,02 * (DO663) + 20,20 * (DO645). Активность кислотной фосфатазыв корнях пшеницы
КорниАктивность АФазы измеряли согласно Bargaz et al.(2017). Образцы свежей массы корней (100 мг) измельчали с экстракционной смесью, состоящей из 500 мкл 0,1 М буфера ацетата натрия (pH 5,6), содержащего 1 мМ дитиотреитол. Гомогенаты центрифугировали (13000 г при 4 ° C в течение 30 мин) и супернатант (50 мкл) использовали для количественного определения активности APase корня. p -нитрофенилфосфат ( p NPP) использовали в качестве субстрата, и активность фермента определяли как количество, гидролизующее 1 нмоль p NPP в минуту на 1 г свежей массы корней.
Статистический анализ
Статистический анализ данных проводился с помощью программного обеспечения IBM ® SPSS ® Statistics V. 24. Использовали однофакторный дисперсионный анализ (дисперсионный анализ) с последующим апостериорным тестом Тьюки для определения значимой разницы между средствами обработки при уровне значимости 0,05. Анализ PCA выполняли с использованием статистической программы Minitab V.18.
Результаты
Биохимические свойства PSB и влияние на рост корней проростков пшеницы
Идентификация PSB и
In vitro свойствНа основании секвенирования гена 16S рРНК, изоляты PSB, использованные в этом исследовании, принадлежат к родам Pseudomonas .Изоляты PSB имели разные PSC от TCP в диапазоне от 113 до 121,2 мг P l -1 для PSB 3 и PSB 5 (высокий PSC), от 88,79 до 99,88 мг л -1 для PSB 2 и PSB 4 (средний PSC) и до 41,37 мг P l -1 для PSB 1 (низкий PSC) (таблица 1). Ясные ореолы, солюбилизирующие P, вокруг бактериальных колоний наблюдались у всех изолятов и варьировали от 4,9 до 5,8. Подкисление среды с помощью RP или TCP резко упало с начального значения 7 до 4.34, за исключением изолята PSB 1 , pH среды которого поддерживали около нейтрального в течение пяти дней инкубации. Кроме того, PSB были оценены как изоляты, продуцирующие ИУК (10,46–36,41 мкг мл −1 ), N 2 -фиксаторы (производство аммиака от 0,02 до 0,19 мкмоль мл −1 ), продуценты сидерофоров, HCN производители, а также солеустойчивое выращивание до 0,86 М NaCl.
Таблица 1 Свойства солюбилизирующих фосфат бактериальных изолятов (PSB 1 до PSB 5 ), связанные с солюбилизацией P, индексом солюбилизации в чашке с агаром, доступным P в инокулированной почве (ppm), подкислением среды, продуцированием IAA, аммиаком продукция, индекс сидерофоров, продукция HCN и толерантность к солености.
PSB
In vivo Влияние на рост корней проростков пшеницыИзмерение морфологических признаков роста корней как у 7-, так и у 15-дневных инокулированных проростков показало значительное увеличение большинства признаков корешка, за исключением, отмеченного для RD, не показавшего никаких различий, поскольку по сравнению с неинокулированными сеянцами (таблица 2). В частности, PSB 1 и PSB 3 значительно улучшили RL (34,40%), RSA (34,04%) и RV (32,5%) 7-дневных проростков; Между тем, именно PSB 4 значительно увеличил RL (58.54%), RSA (65,55%) и RV (77,77%) 15-дневных проростков. Этот изолят имел наибольший эффект на 15-дневные проростки, особенно на RSA и RV, который значительно увеличился на 126,33 и 60% по сравнению с 7-дневными сеянцами.
Таблица 2 Вариации морфологических признаков корней в возрасте 7, 15 и 42 дней твердой пшеницы, удобренной камнем P, в ответ на инокуляцию пятью изолятами PSB по сравнению с обработкой только P (RP и TSP).
Влияние изолятов PSB на растения твердой пшеницы в возрасте 42 дней, снабженных RP
Влияние на параметры роста растений пшеницы 3).Для всех изолятов PSB этот положительный эффект был значительным по сравнению с растениями пшеницы, удобренными только RP. PSB
2 значительно увеличил как SDW, так и RDW по сравнению с растениями пшеницы, удобренными RP или TSP. Это увеличение по PSB 2 показало самый высокий SDW по сравнению с RP (48%), а не TSP (34%). Также были отмечены различия между изолятами PSB в отношении RDW (хотя и незначительные), в частности, для PSB 5 и PSB 4 , показывающих наивысший и самый низкий RDW, соответственно.Таблица 3 Вариации роста твердой пшеницы, удобренной камнем P, в ответ на инокуляцию пятью изолятами PSB по сравнению с обработкой только P (RP и TSP).
Влияние на морфологические признаки корней пшеницы
Морфологические признаки корня (например, RL, RSA, RV, RD, количество кончиков (Ntips), количество скрещиваний (Ncross) и количество лягушек (Nfroks)) у 42-дневного возраста Инокулированные растения заметно улучшились по сравнению с неинокулированными растениями (Таблица 2). Были отмечены очевидные различия между изолятами PSB, особенно PSB 2 , чье влияние на корневые признаки (за исключением RD) оказалось наиболее значительным по сравнению с обоими RP- и TSP-удобренными и неинокулированными растениями.Аналогичные эффекты были отмечены для остальных изолятов PSB, но в меньшей степени, чем для PSB 2 . Значительные различия были обнаружены с PSB 3 и PSB 4 , в частности, значительное увеличение RL (на 37 и 34%) и RSA (на 66,57 и 53,56%) по сравнению с применением RP, а не TSP. Кроме того, оба изолята «PSB 1 и PSB 5 » также положительно влияли на свойства корней пшеницы, хотя различия остаются незначительными по сравнению с растениями пшеницы, удобренными RP или TSP.
Кроме того, конкретные признаки корня, такие как удельная площадь корня (SRA) и удельная длина корня (SRL), выявили значительные различия в ответе на инокуляцию PSB (рис. 1). И SRL, и SRA были самыми высокими у пшеницы, инокулированной PSB 4 , по сравнению с применением RP (увеличение на 59 и 56%) и TSP (увеличение на 33 и 30%). Остальные изоляты PSB также показали аналогичные тенденции для SRL и SRA по сравнению с применением RP, а не TSP, хотя и в меньшей степени, чем PSB 4 .
Рисунок 1 Вариации удельной длины корня (A) и удельной площади корня (B) твердой пшеницы, удобренной камнем P, в ответ на инокуляцию пятью изолятами PSB по сравнению с обработками P (RP и TSP) только через 42 дня после прорастания. Планки погрешностей представляют собой стандартное отклонение для четырех повторностей, и каждая повторность состоит из восьми растений пшеницы на горшок. Средние значения, отмеченные одной и той же буквой, существенно не различались при p <0.05 .
Влияние изолятов PSB на надземные и подземные физиологические модификации твердой пшеницы, полученной с RP
Влияние на питание P пшеницы, полученной с RP
Содержание фосфора в побегах и корнях увеличивалось в ответ на инокуляцию PSB по сравнению с необработанными. – инокулированные растения как у 15-, так и у 42-дневных растений пшеницы (таблица 4). Для содержания Pi как в побегах, так и в корнях значительное увеличение было отмечено в ответ на инокуляцию PSB 1 и PSB 2 (и PSB 5 для Pi побега) по сравнению с RP-, но также и с TSP-удобрением. растения с точки зрения корня Pi.Эта тенденция имела тенденцию изменяться у 42-дневных растений пшеницы, у которых вариации содержания Pi были наиболее значительными в побегах (самые высокие) и корнях (самые низкие) в ответ на инокуляцию PSB 3 по сравнению с растениями, удобренными RP. Однако инокуляция PSB 1 индуцировала (42-дневные растения) значительное накопление Pi в корнях по сравнению с растениями, удобренными RP или TSP. Что касается общего содержания фосфора в побегах у 42-дневных растений, PSB 5 индуцировал более высокое накопление по сравнению со всеми инокулированными и неинокулированными обработками.Наоборот, накопление Pi в корнях значительно ниже, чем у растений, удобренных TSP.
Таблица 4 Вариации содержания фосфора (общего и неорганического) как в корнях, так и в побегах, доступном фосфоре в субстрате для роста ризосферы и активности АРазы в корнях на двух стадиях роста растений пшеницы, удобренных камнем P и инокулированных пятью изолятами PSB по сравнению с Только лечение P (RP и TSP).
Влияние на доступный в ризосфере P и активность корней APase пшеницы
Результаты в таблице 4 показывают, что доступный P в ризосфере увеличился во всех инокулированных RP-оплодотворенных 15-дневных проростках, хотя и незначительно ни между изолятами, ни обработками TSP.Однако этот параметр значительно снизился у 42-дневных растений, инокулированных PSB 1 , PSB 3 и PSB 5 , которые показали лучшее усвоение P корнями, поскольку это подтверждалось более высоким содержанием P в побегах по сравнению с RP- удобренные растения (таблица 4). Однако растения, удобренные TSP, показали самую высокую доступную в ризосфере фракцию P по сравнению со всеми обработками. Кроме того, P-гидролизирующая APase в корнях пшеницы варьировала в ответ на инокуляцию и стадию роста растений, особенно у 42-дневных растений пшеницы, у которых активность APase корня значительно увеличивалась вместе со снижением доступности P в ризосфере (Таблица 4).Эта тенденция была в основном отмечена в корнях, инокулированных PSB 1 , PSB 2 и PSB 5 , чья активность APase была почти в пять раз выше, чем у неинокулированных растений пшеницы, удобренных RP и TSP.
Влияние на эффективность сбора фосфора корня
У растений, удобренных TSP, самая высокая RPAE по сравнению с самой низкой эффективностью у растений, удобренных RP (рис. 2A). Инокуляция растений, удобренных RP, PSB 1 , PSB 3 , PSB 4 и PSB 5 не повлияла на RPAE, вариации которых остаются незначительными по сравнению с таковыми у растений, удобренных RP.Только PSB 2 значительно увеличивал (129,59%) RPAE по сравнению с неинокулированными растениями, удобренными RP. Такое заметное увеличение также было значительно выше по сравнению с остальными изолятами PSB, но в меньшей степени с PSB 4 . С другой стороны, инокулированные растения демонстрировали очень значимую и положительную корреляцию (R = 0,6, p = p = 0,0014 ** ) между содержанием неорганического «Pi» и общим содержанием P, что указывает на то, что изоляты PSB, вероятно, способствуют лучшая эффективность внутреннего использования фосфора (рис. 2В).
Рисунок 2 Вариация эффективности приобретения корневого P RPAE (A) и взаимосвязь между неорганическим P и общим P твердой пшеницы (B) , удобренной каменной P, в ответ на инокуляцию пятью изолятами PSB по сравнению с Только лечение P (RP и TSP). Планки погрешностей представляют собой стандартное отклонение четырех повторов, каждый из которых состоит из восьми растений пшеницы на горшок, собранных через 42 дня после прорастания. Средние значения, отмеченные одной и той же буквой, существенно не различались при p <0.05 . На панели (B) черные кружки: растения, удобренные RP, инокулированные пятью изолятами PSB, состоящими из двадцати повторов (четыре повтора на PSB). Серые квадраты: удобренные RP и TSP растения, состоящие из четырех повторностей, каждая повторность состоит из восьми растений пшеницы на горшок.
Влияние на содержание хлорофилла (a и b) и проводимость в устье , В
) .Только инокуляция PSB 1 и PSB 5 значительно улучшила содержание Chl b по сравнению с растениями, удобренными TSP и RP (рис. 3A). Точно так же только PSB 1 значительно увеличил содержание Chl a и показал, среди всех изолятов PSB, самое высокое содержание Chl a по сравнению с контролями (рис. 3B). Следует отметить, что PSB 2 и PSB 3 также значительно повысили содержание Chl a по сравнению с неинокулированными растениями, удобренными только RP. Аналогичным образом, устьичная проводимость (gs) была значительно выше у инокулированных растений пшеницы независимо от изолятов PSB со средним увеличением на 71.71 и 58,62% по сравнению с растениями пшеницы, удобренными RP и TSP, соответственно (рис. 3C).Рисунок 3 Вариации содержания хлорофилла b в листьях (A) , содержания хлорофилла a в листьях (B) и устьичной проводимости (gs) (C) твердой пшеницы, удобренной камнем P, в ответ на инокуляцию с пятью изолятами PSB только против P (RP и TSP). Планки погрешностей представляют собой стандартное отклонение для четырех повторностей, и каждая повторность состоит из восьми растений пшеницы на горшок.Средние значения, обозначенные одной и той же буквой, значимо не различались при p <0,05 .
Значительные корреляции были обнаружены между общим содержанием Chl и содержанием как Pi (R = 0,5, p = 0,001 **), так и общего P (R = 0,6, p = 0,001 **) инокулированной пшеницы, удобренной RP растения (Рисунки 4A, B ) . Такие положительные корреляции могут относиться как к использованию, так и к физиологической эффективности RP для лучшей активности фотосинтеза у инокулированных растений, чем у неинокулированных растений.Эта положительная взаимозависимость может быть оценена до 36 и 22% на основе наклона регрессионной модели (y = ax + b) содержания Chl как функции Pi и общего содержания P в растении, соответственно.
Рис. 4 Корреляция между общим содержанием хлорофилла и растительным P (неорганическое (A) и общее (B) содержание в твердой пшенице, инокулированной пятью изолятами PSB и удобренной камнем P (черные кружки) против без инокулированная пшеница, удобренная только обработкой P (RP и TSP) (серые квадраты).Черные кружки: растения, удобренные RP, инокулированные пятью изолятами PSB, состоящими из четырех повторов на PSB. Серые квадраты: растения, удобренные RP и TSP, состоящие из трех повторов на RP и четырех повторов на TSP. Для каждой обработки каждая повторность состояла из восьми растений пшеницы на горшок.
Влияние на содержание азота и белка в побегах твердой пшеницы
Растения пшеницы Содержание азота повышается в ответ на инокуляцию PSB со средним увеличением на 10,04% по сравнению с неинокулированными растениями, удобренными только RP (таблица 5), хотя существенной разницы нет. ( P <0.05 ) отмечен среди инокулированных и неинокулированных растений. Однако удобрение TSP значительно увеличило содержание N на 19 и 30,66% по сравнению с инокуляцией PSB и удобрением RP, соответственно. С другой стороны, хотя содержание N в инокулированных растениях не увеличилось значительно по сравнению с растениями, удобренными RP, NUE выявил значительные эффекты из-за инокуляции PSB 3 (78%), PSB 5 (50%) и PSB . 1 (37,5%) по сравнению с TSP (самое высокое содержание N) и удобрением RP.Подобно содержанию N, растения, удобренные TSP, имели самое высокое содержание белка по сравнению со всеми обработками (Таблица 5). Тем не менее, по сравнению с неинокулированными растениями, статистически большее количество белка было увеличено в ответ на PSB 2 , PSB 3 , PSB 5 и PSB 4 на 134,75, 131,52, 57,60 и 40,76%. соответственно.
Таблица 5 Вариации содержания азота и белка в твердой пшенице, удобренной камнем P, в ответ на инокуляцию пятью изолятами PSB по сравнению с обработкой только P (RP и TSP).
Влияние инокуляции PSB на взаимозависимость между надземными и подземными признаками
Многопараметрический корреляционный анализ между всеми надземными (например, биомасса, физиологические признаки, содержание фосфора и т. Д.) И подземными (например, корневыми признаками) (APase корня, доступный P)) выявили существенные различия между инокулированными и неинокулированными растениями (фиг. 5; дополнительная таблица S2). Анализ главных компонентов показал, что сгруппированные группы, в которых имеются определенные корневые черты (например,грамм. SRL, SRA, RDW, Ntips) были тесно связаны с содержанием N и P как в побегах, так и в корнях инокулированных растений пшеницы (рис. 5A), что также подтверждается в корреляционной матрице (дополнительная таблица S2), показывающей дифференциальные ответы между изолятами PSB. Более того, инокулированные растения представляли вторую группу параметров кластеризации, связанных с морфологическими признаками корня (например, RL, RV, RD, Ncross и NForks), физиологическими признаками (например, содержанием белка и N, gs) и доступным P в ризосфере, которые все указывают на важность морфологических признаков корней в поглощении корнями питательных веществ и абсорбционной способности (рисунки 2 и 5A; дополнительная таблица S2).Кроме того, инокуляция изолятами PSB, по-видимому, имеет положительную взаимозависимость между общим Chl, содержанием Pi в корне, SDW и корневой APase. В отличие от инокулированных растений, у неинокулированных (оплодотворенных RP или TSP) морфологические признаки корней (например, RV, SRL, RSA и Nfroks) коррелировали с содержанием P в побегах, но без корреляции между морфологическими признаками корня, активностью APase, содержимое корневого Pi или даже RDW, которое сгруппировано отдельно (рис. 5B). Более того, вторая группа, объединяющая физиологические параметры и параметры роста (содержание Хл, ВСП, содержание белка), имела положительную корреляцию с наличием фосфора в почве в ризосфере.Также было отмечено, что большинство функциональных признаков, участвующих в поглощении P (например, RDW, доступная P, APase и признаки корня, разбросаны в обратном направлении как морфологических, так и физиологических признаках корня (например, содержание Chl, белка и N, gs), что правдоподобно указывает на несбалансированное использование этих признаков, ответственных за приобретение фосфора и ростовые показатели.
Рисунок 5 Анализ основных компонентов, разработанный на основе роста, свойств корней и физиологических параметров, измеренных у твердой пшеницы, удобренной камнем P, в ответ на инокуляцию пятью изолятами PSB по сравнению с неинокулированных растений, удобренных только обработкой P (RP и TSP).Данные представляют собой средние значения четырех повторностей, и каждая повторность состоит из восьми растений пшеницы на горшок, собранных через 42 дня после прорастания. Pt отросток, всего P отростка; Пи отросток, внутриклеточный P отростка; Pt root, всего P корня; Корень Pi, внутриклеточный P корня; N – поглощение азота; P Olsen, P в ризосфере; APase, активность кислой фосфатазы в корне; Chl b – содержание хлорофилла b в листьях; Chl a, содержание хлорофилла a в листьях; Chl tot, общий хлорофилл; gs – устьичная проводимость; Prot, содержание белка; RDW – сухой вес корня; SDW, снимать сухой вес; SRL, удельная длина корня; SRA, специфическая корневая зона; RL – длина корня; RSA, площадь поверхности корня; RD, диаметр корня; RV, корневой объем; Ncross, количество переходов; NTips, количество подсказок; Nforks, Кол-во вил.
Обсуждение
Настоящее исследование вносит свой вклад в имеющиеся знания о взаимодействии растений пшеницы и PSB при низкой доступности фосфора, в частности, об изменениях морфологических признаков корней вместе с соответствующими модификациями ризосферы и надземных физиологических параметрах, связанных с эффективностью использования фосфора. Мы демонстрируем значительное влияние пяти изолятов PSB, контрастирующих по их PSC, на доступность P в ризосфере, морфологические черты корня и улучшенные надземные параметры, вариации которых могут помочь в понимании очень сложных взаимодействий корня и PSB при низких доступных формах P, таких как порода P.Мы также рассматриваем гипотезу о том, что PSB может иметь более сильное влияние на биофизические характеристики корней (больше, чем локализованная солюбилизация P ризосферы), положительно отражаясь на способности корней усваивать питательные вещества (включая P среди прочего) и общую физиологию урожая. Кроме того, наши результаты предполагают, что бактерия с высоким уровнем растворения в планшете P не обязательно указывает на важные ответы in-planta , учитывая, что временные модификации на подземном уровне оказались специфичными для PSB, независимо от способности солюбилизировать P они были впервые выбраны для.
P Солюбилизирующие ризобактерии по-разному влияли на развитие проростков твердой пшеницы
Было продемонстрировано, что способность изолятов PSB солюбилизировать две разные формы P (например, RP и TCP) положительно влияет на развитие проростков пшеницы, улучшение, которое может быть связано не только с повышенная биосолюбилизация ризосферы P и модификации морфологии корня (таблица 2; дополнительная таблица S1), но также и множественные признаки PGP (например, IAA, сидерофор, NH 4 + и т. д.); Таблица 1), вероятно, способствует дополнительному улучшению роста. Улучшение морфологических характеристик корней проростков в ответ на инокуляцию (в частности, PSB 1 и PSB 4 ), вероятно, было следствием более высокой продукции ИУК, гормона-регулятора роста растений, который, помимо ускорения роста и удлинения корней, также улучшает фотосинтетическую способность, углеводный обмен и общий урожай растений (Li et al., 2019). В этом контексте недавнее исследование Marathe et al.(2017) продемонстрировали способность продуцирующего ИУК штамма PSB ( Pseudomonas aeruginosa ) стимулировать прорастание семян на 2 дня раньше и увеличивать как поглощение питательных веществ (N, P и K), так и содержание хлорофилла (chl a и chl b). Это полностью согласуется с выводами текущего исследования, а также с рядом предыдущих исследований (Khiangte and Lalfakzuala, 2011; Linu et al., 2019; Liu et al., 2019), хотя для определения любого из них потребуется точный количественный анализ. количество и тип ИУК, которые, вероятно, ответственны за рост корней (Kowalczyk and Sandberg, 2001; Liu et al., 2012). Однако остается неясным, почему инокуляция PSB 3 , которая показывает самую высокую скорость продуцирования ИУК, не повлияла на свойства корня (особенно у 15-дневных проростков) по сравнению с остальными изолятами PSB или неинокулированными проростками. Другие бактериальные признаки, такие как выработка аммиака, подкисление среды и устойчивость к осмотическому стрессу (таблица 1), могут положительно повлиять на устойчивость проростков, включая важную способность корня к абсорбции питательных веществ на ранней стадии, которая может улучшить солюбилизацию и последующее использование RP (Abbasi et al., 2015; Кумар, 2016; Перес и др., 2016).
Бактериальные признаки PGP хорошо задокументированы (Cerozi and Fitzsimmons, 2016; Vandamme et al., 2016; Paul and Sinha, 2017; Suleman et al., 2018; Liu et al., 2019; Parastesh et al., 2019) в то же время с точки зрения увеличения роста и урожайности растений пространственные и временные изменения функциональных характеристик корней в ответ на инокуляцию PSB все еще малоизвестны. В этом исследовании модификации ризосферы на ранней стадии развития растений (7- и 15-дневные проростки), вероятно, связаны с инокуляцией изолятов PSB, скорость растворения P которых в почве была почти в три раза выше, чем у неинокулированных растений без растений. почвы, что согласуется с данными в пластине (таблица 1).Кроме того, вариации в отношении солюбилизации P как in vitro , так и в почве без растений указали на явные различия между изолятами PSB, особенно с PSB 1 , которые значительно увеличили RL и RSA у 7-дневных проростков, демонстрируя при этом самые низкие значения. PSC. Такая стимуляция наблюдалась и у 15-дневных проростков, у которых содержание фосфора (особенно в корнях) значительно увеличивалось в ответ на инокуляцию PSB 1 (таблица 2). Напротив, изолят PSB 5 , показывающий самый высокий PSC, не оказывал значительного влияния на 7-дневные проростки пшеницы по сравнению с неинокулированными проростками, снабженными RP.Более того, наблюдаемые вариации у 15-дневных проростков, по-видимому, были специфичными для PSB, учитывая, что только PSB 4 (умеренный PSC) вызывал значительные улучшения корневых признаков, в основном RL, RSA и RV (Таблица 2). Такие вариации ризосферы на ранней стадии, которые впервые были развернуты в настоящем исследовании, могут указывать на явные дифференциальные эффекты, которые PSB может оказывать на проростки пшеницы на ранней стадии роста. Это приведет к возникновению новых исследовательских вопросов, позволяющих лучше понять возможные способы действий, которые может развиваться интерфейс PSB – корень, а не рутинно оцениваемая солюбилизация бактериального P, и в настоящее время считается, что это имеет решающее значение для скрининга эффективных PSB.
P Солюбилизирующие бактерии могут иметь большее влияние на корневую систему, чем ризосфера P Солюбилизация
Помимо прямого воздействия, которое PSB может оказывать на солюбилизацию RP, кажется, что со временем меняются морфологические признаки корня (например, 7-, 15- и 42 стародневные растения пшеницы) были связаны с высшим ростом растений (таблицы 2 – 4). После 15 дней роста инокуляция всеми изолятами PSB значительно улучшила RDW, тогда как доступный в ризосфере P существенно не изменился, даже несмотря на то, что может наблюдаться небольшое увеличение по сравнению с растениями, удобренными RP (таблица 4).Корни 42-дневных растений пшеницы также показали аналогичную тенденцию при инокулировании PSB 1 , PSB 2 и PSB 5 . Для растений, инокулированных PSB 5 (демонстрирующих самый высокий PSC), меньшая доступная фракция P в ризосфере может частично объясняться важной RDW и глубиной корня, которые указывают на важный рост корней, предположительно ответственный за повышенное поглощение P корнями и содержание P в побегах пшеницы. (Таблица 4). В соответствии с этими выводами имеются экспериментальные данные о биосолюбилизации почвенного бактериального фосфора (Mamta et al., 2010; Батул и Икбал, 2019; Emami et al., 2019), между тем взаимосвязь между PSB и функциональными признаками корня и их распространение в ризосфере еще не полностью расшифрована, и тем более, когда речь идет, в частности, о временных вариациях на разных стадиях роста сельскохозяйственных культур. В целом, наши результаты впервые показывают, что PSB может демонстрировать различные PSC во время роста (три стадии роста растений) и что скорость солюбилизации P на уровне ризосферы, вероятно, трудно оценить, в то время как корни продолжают расти в пространстве и во времени.Можно предположить, что точный скрининг PSB определенно будет учитывать как биохимические особенности PSB, так и реакции взаимодействия корневых бактерий, что свидетельствует о том, что ассимиляция Р ризосферы является биологическим процессом, управляемым корнями, который в значительной степени зависит от общих характеристик подземного роста.
Насколько нам известно, и даже несмотря на то, что параметры корней были измерены деструктивно, это исследование является первым, описывающим интересные временные вариации как морфологических, так и физиологических характеристик корней пшеницы в ответ на инокуляцию PSB, которые контрастируют с их PSC.Действительно, наши результаты подтверждаются большинством исследований, основанных на PSB и / или PGPR, в которых реакции роста культур, особенно модификации корней, измерялись либо на ранних (прорастание и всходы), либо на поздних стадиях сбора урожая без учета контраста, который PSB может иметь на P степень солюбилизации, которую они могут выражать на разных стадиях развития растений. В целом, на трех стадиях роста растений (7-, 15- и 42-дневный) морфологические характеристики корней (например, объем, поверхность, диаметр и количество кончиков и вилок), очевидно, увеличились в ответ на инокуляцию, которая позволила эффективно использовать RP , что согласуется с недавними выводами Ванга и Чу (2015); Suleman et al.(2018); Лю и др. (2019) и Wang et al. (2019). Эти авторы продемонстрировали, что помимо солюбилизации фосфора, инокуляция PSB может изменять функционирование корней за счет модуляции экспрессии ауксин-чувствительных генов, тем самым играя важную роль в регуляции уровня эндогенной ИУК с положительными последствиями для приобретения фосфора и физиологического статуса растений. Кроме того, пространственная неоднородность ризосферы / корня может возникать из-за более интенсивного изучения почвы, что приводит к более высокой солюбилизации и поглощению фосфора корнями, что может быть следствием продуцирующих AIA изолятов PSB (в частности, PSB 1 , PSB 2 и PSB ). 3 ), вероятно, участвует в регуляции морфологии корневой системы, такой как удлинение боковых корней (Raya-Gonzalez et al., 2014). Более того, сильные и эффективные системы укоренения у инокулированных растений пшеницы были связаны с более высоким содержанием азота в побегах и листового белка, особенно в ответ на PSB 2 , характеризуемый как производящий аммоний (эквивалентный 80 нмоль мл -1 ) изолят и вероятно, участвует в несимбиотической фиксации N 2 во время роста растений. Корреляционный анализ (дополнительная таблица S2) также показал тесную взаимосвязь между N побегов, содержанием белка в листьях и характеристиками корней (например,грамм. RSA, RD и N наконечники), что свидетельствует о более сильном подземном эффекте. В частности, морфологические признаки корня (например, SRL, SRA, RDW, Ntips) могут в значительной степени способствовать лучшему усвоению как N, так и P (Рисунок 5; Дополнительная таблица S2). Однако необходимы продвинутые мультидисциплинарные подходы, в частности, комбинирование методов фиксации N 2 (т.е. естественное изобилие 15 N), выделение подземных фотосинтатов и заселение корнями внесенных кандидатов PSB. В этом контексте было подсчитано, что до 60% фотосинтезированного углерода в пшенице, горохе, кукурузе и томатах перемещается под землю, где связанные с корнями микроорганизмы могут его метаболизировать или использовать для роста растений и роста растений. микробиом ризосферы (Morgan et al., 2005; Эрнандес и др., 2015; Wang et al., 2016).
Ризосферная бактериальная биосолюбилизация фосфора предположительно стимулирует положительные взаимодействия над и под землей
Большинство предыдущих исследований в значительной степени описывали прямое влияние PSB на рост растений, основанное, главным образом, на солюбилизации фосфора и росте растений (Manzoor et al. ., 2017; Singh et al., 2018; Liu et al., 2019). В других исследованиях также сообщалось о важных положительных эффектах на несколько параметров корня (Sarsan, 2016; Suleman et al., 2018; Rezakhani et al., 2019) без сильной связи с надземными P-параметрами. Тем не менее, это исследование предоставило новые доказательства того, что эффекты PSB в основном не ограничиваются только солюбилизацией P, а распространяются на множество известных и неизвестных косвенных эффектов на морфологические признаки корня, тем самым улучшая как приобретение, так и внутреннее использование P. Множественные дифференциальные ответы на уровне Взаимодействия «PSB-растение» описаны здесь, в частности, о значительном увеличении доступности фосфора в ризосфере из-за инокуляции PSB (таблица 1) на ранней стадии развития растения (7-дневные проростки), которые могут легко обеспечить адекватное количество фосфора. доступны для непрерывно растущих корней, таким образом, вероятно, обеспечивается лучший рост растений и питание P для последующих стадий роста.Другой ответ заключается в том, что PSB может играть ключевую роль в росте растений, способствуя развитию корней у 15-дневных проростков в большей степени, чем солюбилизация P, которая, по-видимому, была выражена раньше, на 7-й день после прорастания, и имела тенденцию к снижению в 15-дневный и 42-дневные растения пшеницы. Это согласуется с наблюдением, что в инокулированной почве без растений (результаты не показаны) солюбилизация P была выше после 7 дней инкубации и имела тенденцию к снижению на 15 день с последующим важным (хотя и не значительным) восстановлением солюбилизации P на 42-й день. день.
У растений пшеницы, инокулированных изолятами PSB, большинство надземных и подземных параметров были сгруппированы в одну группу, в частности морфологические признаки корня, такие как Ntips, указывающие на пролиферацию корней (Harmer, 1990), которые в значительной степени коррелировали с содержанием P в корнях и побегах. Эта взаимосвязь может быть объяснена способностью корня к интенсивному кормлению почвы, в том числе благодаря быстрому размножению корней, ведущему к большему поглощению фосфора. Кроме того, положительная корреляция между доступным в ризосфере P и корневыми Ntips и RD в ответ на инокуляцию может указывать на то, что повышенная доступность P в почве ризосферы и его лучшая надземная транслокация происходили прямо или косвенно в ответ на инокуляцию PSB, предположительно из-за лучшее использование морфологических признаков корней (таблица 2), что позволило использовать большую поверхность почвы (более высокие RL, RSA, Nforks, а также удельная длина и площадь корня).Более того, у 42-дневных растений PSB имел тенденцию стимулировать как корневую APase-активность, так и RL в большей степени, чем активность солюбилизации P, которая со временем снижалась из-за увеличения абсорбционной способности корней, а также из-за возможной внутренней ремобилизации клеточных пулов P из-за увеличения корневой Активность АПазы. Это согласуется с предыдущими исследованиями, в которых сообщается о способности штаммов PSB продуцировать APase для улучшения питания P (Behera et al., 2017; Chawngthu et al., 2020). Однако этот признак никогда не отслеживался своевременно в инокулированных корнях или должным образом не рассматривался в качестве важного корневого признака, заслуживающего изучения на этапах скрининга in planta PSB.
Более того, инокулированные растения продемонстрировали явное улучшение надземных физиологических характеристик, примером чего является более высокое содержание P, N и хлорофилла, а также устьичная проводимость по сравнению с неинокулированными растениями. Фотосинтетическая активность (общий Хл) когерентно улучшается с увеличением содержания P в побегах (как общего, так и неорганического) при введении RP и инокуляции PSB (особенно PSB 3 ) (Рисунок 4; Дополнительная таблица S2), что четко объясняется положительными эффектами инокуляции. на разрастание корней, что позволяет получать больше фосфора из почвы, что хорошо коррелирует с содержанием фотосинтетического хлорофилла.Такой вывод указывает на четкие взаимосвязи, которые, вероятно, вызваны PSB при низкой доступности фосфора, по сравнению с адекватным (TSP) минеральным питанием, которое не вызывало такой реакции. Это согласуется с недавними открытиями Wang et al. (2015) и Rozier et al. (2019), что PGPR (например, Azospirillum lipoferum , Azospirillum brasilense и Burkholderia phytofirmans , в частности, фиксация N 2 ) способствовали лучшей фотосинтетической активности кукурузы, пшеницы и просо, однако ни над землей, ни под землей. механистические взаимодействия, связанные с P и PSB, были выделены до сих пор.
Выводы
Хотя исследования, посвященные ОВО, продвинули понимание сложности задействованных механизмов, остается неясным, как ОВО способствует взаимодействию под землей и над землей. Совершенно реально, что PSB вносит непосредственный вклад в солюбилизацию P ризосферы, однако степень, в которой PSB может способствовать биофизическим свойствам корня и наземным физиологическим вариациям, остается загадкой, особенно из-за сложных взаимодействий корня и PSB, которые могут происходить и, вероятно, изменяться с течением времени.Наши результаты показывают, что контраст с точки зрения скорости солюбилизации бактериального P может не быть единственным критерием для различения на ранней стадии ( в скрининге на планшете ) низкоскоростных изолятов PSB, эффект которых на planta может быть значительным из-за специфических взаимодействий с корнями. которые так или иначе позволяют положительные наземные ответы (случай PSB 1 и PSB 4 ). Также можно предположить, что улавливание эффективных PSB и оценка их в planta , способствующих росту признаков с помощью множественных экспериментов по инокуляции, по-прежнему неизбежны, однако реакцию растений следует отслеживать своевременно и в пространстве, чтобы указать «во времени и в пространстве». модификации, позволяющие точно интерпретировать бактериальные эффекты.Такая стратегия, хотя и требует много времени, может предоставить исчерпывающие данные о соответствующей смеси уловки и бактерий «PSB» с дополнительными и синергетическими эффектами на всех стадиях роста растений. Это могло бы существенно дополнить стандартный подход к созданию бактериального консорциума « в планшете », который не учитывает своевременные ответы растений в процессе скрининга. Можно также сделать вывод, что принятие междисциплинарных подходов, особенно фенотипирования функциональных признаков как подземных, так и наземных, вероятно, необходимо для объективной интерпретации результатов и точного скрининга, что в конечном итоге приведет к успешному применению в полевых приложениях .
Заявление о доступности данных
Все наборы данных, созданные для этого исследования, включены в статью / Дополнительные материалы.
Вклад авторов
Все авторы внесли свой вклад в статью и одобрили представленную версию.
Финансирование
Эта работа была поддержана OCP Group – Situation Innovation Group в рамках проекта AS17 (2019-2021), предоставленного AB Политехнического университета Мохаммеда VI (UM6P).
Конфликт интересов
Авторы заявляют, что исследование проводилось в отсутствие каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Благодарности
Мы благодарим профессора Укаррума Абдаллаха за ценную практическую поддержку и советы при анализе данных, связанных с фотосинтезом. Мы благодарны г-ну Рашиду Гани за его доступность и техническую помощь. Авторы искренне благодарят профессора Исмаила Раки (языковая лаборатория UM6P) за ценную помощь в языковом редактировании рукописи.
Дополнительные материалы
Дополнительные материалы к этой статье можно найти в Интернете по адресу: https: // www.frontiersin.org/articles/10.3389/fpls.2020.00979/full#supplementary-material
Ссылки
Аббаси, М. К., Муса, Н., Манзур, М. (2015). Минерализация растворимых фосфорных удобрений и нерастворимого фосфата в ответ на фосфат-солюбилизирующие бактерии и птичий помет и их влияние на рост и эффективность использования фосфора перца чили ( Capsicum annuum L .). Биогеонауки 12, 4607–4619. doi: 10.5194 / bg-12-4607-2015
CrossRef Полный текст | Google Scholar
Аднан, М., Шах, З., Фахад, С., Ариф, М., Алам, М., Хан, И. А. и др. (2017). Фосфатосолюбилизирующие бактерии сводят на нет антагонистический эффект известкования почвы на биодоступность фосфора в щелочных почвах. Sci. Реп. 7, 1–13. doi: 10.1038 / s41598-017-16537-5
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Alzoubi, M. M., Gaibore, M. (2012). Влияние солюбилизирующих фосфат бактерий и органических удобрений на доступность сирийского каменного фосфата и повышение эффективности тройного суперфосфата. World J. Agric. Sci. 8, 473–478. doi: 10.5829 / idosi.wjas.2012.8.5.1668
CrossRef Полный текст | Google Scholar
Bakhshandeh, E., Rahimian, H., Pirdashti, H., Nematzadeh, G.A. (2015). Оценка фосфат-солюбилизирующих бактерий на рост и урожайность зерна риса ( Oryza sativa L .), Возделываемого в северном Иране. J. Appl. Microbiol. 119, 1371–1382. doi: 10.1111 / jam.12938
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Баргаз, А., Ghoulam, C., Amenc, L., Lazali, M., Faghir, M., Abadie, J., et al. (2012). Транскрипт фосфоенолпируватфосфатазы индуцируется в коре корневых клубеньков Phaseolus vulgaris в условиях дефицита фосфора. J. Exp. Бот. 63, 4723–4730. doi: 10.1093 / jxb / ers151
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Bargaz, A., Noyce, G. L., Carlsson, G., Furze, J. R., Jensen, E.J., Dhiba, D., et al. (2017). Взаимодействие видов улучшает распределение корней, микробное разнообразие и накопление фосфора в промежуточных посевах пшеницы и сои в условиях дефицита фосфора. заявл. Soil Ecol. 120, 179–188. doi: 10.1016 / j.apsoil.2017.08.011
CrossRef Полный текст | Google Scholar
Bargaz, A., Lyamlouli, K., Chtouki, M., Zeroual, Y., Dhiba, D. (2018). Микробные ресурсы почвы для повышения эффективности удобрений в интегрированной системе управления питательными веществами. Фронт. Microbiol. 9, 1–25. doi: 10.3389 / fmicb.2018.01606
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Barra, P. J., Inostroza, N.Г., Акуна, Дж. Дж., Мора, М. Л., Кроули, Д. Э., Хоркера, М. А. (2016). Составление бактериальных консорциумов из авокадо (Persea Americana Mill.) И их влияние на рост, биомассу и активность супероксиддисмутазы проростков пшеницы в условиях солевого стресса. заявл. Soil Ecol. 102, 80–91. doi: 10.1016 / j.apsoil.2016.02.014
CrossRef Полный текст | Google Scholar
Batool, S., Iqbal, A. (2019). Ризобактерии, солюбилизирующие фосфат, как альтернатива химическим удобрениям для роста и урожайности Triticum aestivum (Var.Галактика 2013). Saudi J. Biol. Sci. 26, 1400–1410. doi: 10.1016 / j.sjbs.2018.05.024
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бехера, Б. К., Ядав, Х., Сингх, С., К. Мишра, Р. Р., Сетхи, Б., К. Датта, С. К. и др. (2017). Солюбилизация фосфатов и активность кислой фосфатазы Serratia sp. изолирован из мангровых зарослей дельты реки Маханади, Одиша, Индия. Inter. J. Genet. Англ. Биотех. 15, 169–178. doi: 10.1016 / j.jgeb.2017.01.003
CrossRef Полный текст | Google Scholar
Betencourt, E., Дюпютель, М., Коломб, Б., Дескло, Д., Хинзингер, П. (2012). Пересечение повышает способность твердых сортов пшеницы и нута увеличивать доступность фосфора в ризосфере в почве с низким содержанием фосфора. Soil Biol. Biochem. 46, 181–190. doi: 10.1016 / j.soilbio.2011.11.015
CrossRef Полный текст | Google Scholar
Biswas, J. K., Banerjee, A., Rai, M., Naidu, R., Biswas, B., Vithanage, M., et al. (2018). Возможное применение выбранных металл-устойчивых фосфат-солюбилизирующих бактерий, выделенных из кишечника дождевого червя ( Metaphire posthuma ), для стимулирования роста растений. Geoderma 330, 117–124. doi: 10.1016 / j.geoderma.2018.05.034
CrossRef Полный текст | Google Scholar
Cerozi, B. da S., Fitzsimmons, K. (2016). Влияние pH на доступность фосфора и его состав в питательном растворе для аквапоники. Биоресурсы. Technol. 219, 778–781. doi: 10.1016 / j.biortech.2016.08.079
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Chawngthu, L., Hnamte, R., Lalfakzuala, R. (2020). Выделение и характеристика ризосферных бактерий, солюбилизирующих фосфат, из заболоченных рисовых полей Мизорама, , Индия . Geomicrobiol. J. 37, 366–375. doi: 10.1080 / 014.2019.1709108
CrossRef Полный текст | Google Scholar
Чен, Ю. П., Рекха, П. Д., Арун, А. Б., Шен, Ф. Т., Лай, В. А., Янг, К. С. (2006). Солюбилизирующие фосфат бактерии из субтропической почвы и их способность растворять трикальцийфосфат. заявл. Soil Ecol. 34, 33–41. doi: 10.1016 / j.apsoil.2005.12.002
CrossRef Полный текст | Google Scholar
Де Фрейтас, Дж. Р., Банерджи, М.Р., Гермида, Дж. Дж. (1997). Солюбилизирующие фосфат ризобактерии усиливают рост и урожай, но не поглощают фосфор канолы ( Brassica napus L .). Biol. Fertil. Почвы 24, 358–364. doi: 10.1007 / s003740050
CrossRef Полный текст | Google Scholar
Дель Пилар Лопес-Ортега, М., Криолло-Кампос, П. Дж., Гомес-Варгас, Р. М., Камело-Русинке, М., Эстрада-Бонилья, Г., Гарридо-Рубиано, М. Ф. и др. (2013). Характеристика диазотрофных фосфат-солюбилизирующих бактерий как стимуляторов роста растений кукурузы. Rev. Colomb. Biotecnol. 15, 115–123. doi: 10.15446 / rev.colomb.biote.v15n2.36303
CrossRef Полный текст | Google Scholar
Ditta, A., Imtiaz, M., Mehmood, S., Rizwan, M. S., Mubeen, F., Aziz, O., et al. (2018). Органическое удобрение, обогащенное каменными фосфатами и содержащими фосфат-солюбилизирующие микроорганизмы, улучшает клубенькообразование, рост и урожайность бобовых культур. Commun. Почвоведение. Завод анальный. 49, 2715–2725. doi: 10.1080 / 00103624.2018.1538374
CrossRef Полный текст | Google Scholar
Джаджагло, Д., Рихтер, К. (2008). Эффективность поглощения фосфора растениями Sorghum bicolor (L.) Moench и Phaseolus vulgaris L . Agrosolutions 19, 45–50.
Google Scholar
Эмами, С., Алихани, Х.А., Поурбабаей, А.А., Этесами, Х., Сармадиан, Ф., Мотесшарезаде, Б. (2019). Влияние ризосферных и эндофитных бактерий с множеством признаков, способствующих росту растений, на рост пшеницы. Environ. Sci. Загрязнение. 26, 19804–19813.doi: 10.1007 / s11356-019-05284-x
CrossRef Полный текст | Google Scholar
Fahad, S., Hussain, S., Bano, A., Saud, S., Hassan, S., Shan, D., et al. (2015). Потенциальная роль фитогормонов и ризобактерий, способствующих росту растений, при абиотических стрессах: последствия для изменения окружающей среды. Environ. Sci. Загрязнение. Res. 22, 4907–4921. doi: 10.1007 / s11356-014-3754-2
CrossRef Полный текст | Google Scholar
Fankem, H., Nwaga, D., Deubel, A., Dieng, L., Мербах, В., Этоа, Ф. X. (2006). Возникновение и функционирование фосфатсолюбилизирующих микроорганизмов из ризосферы масличной пальмы ( Elaeis guineensis ) в Камеруне. Afr. J. Biotechnol. 5, 2450–2460. doi: 10.4314 / ajb.v5i24.56044
CrossRef Полный текст | Google Scholar
Фернандес, Л. А., Залба, П., Гомес, М. А., Сагардой, М. А. (2007). Фосфат-солюбилизирующая активность бактериальных штаммов в почве и их влияние на рост сои в тепличных условиях. Biol. Fertil. Почвы 43, 805–809. doi: 10.1007 / s00374-007-0172-3
CrossRef Полный текст | Google Scholar
Gao, X., Shi, D., Lv, A., Wang, S., Yuan, S., Zhou, P. (2016). Повышение доступности фосфора за счет использования сидерата люцерны ( Medicago sativa л) в рисовой агроэкосистеме ( Oryza sativa л). Sci. Реп. 6, 1–13. doi: 10.1038 / srep36981
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Geetha, K., Венкатешам, Э., Хиндумати, А., Бхадраиа, Б. (2014). Выделение, скрининг и характеристика бактерий, способствующих росту растений, и их влияние на Vigna Radita (L.) R.Wilczek. Внутр. J. Curr. Microbiol. Прил. Sci. 3, 799–899.
Google Scholar
Джиро, В. Б., Джиндо, К., Витторацци, К., Де Оливейра, Р. С. С., Консейсао, Г. П., Канеллас, Л. П. и др. (2016). Каменный фосфат в сочетании с фосфат-солюбилизирующими микроорганизмами и гуминовым веществом для снижения потребности растений в фосфоре из простого суперфосфата. Acta Hortic. 1146, 63–68. doi: 10.17660 / ActaHortic.2016.1146.8
CrossRef Полный текст | Google Scholar
Gomes, E. A., Silva, U. C., Marriel, I. E., Oliveira, C. A., Lana, U. G. P. (2014). Микроорганизмы, солюбилизирующие каменный фосфат, выделенные из почвы ризосферы кукурузы. Ред. Бюстгальтеры. Милхо Сорго. 13, 69–81. doi: 10.18512 / 1980-6477 / rbms.v13n1p69-81
CrossRef Полный текст | Google Scholar
Гупта, С., Дангаяч, С., Сундари, С. К. (2015).Изучение роли pgpm в поддержке роста растений в условиях стресса, вызванного фосфорорганическими пестицидами. Глоб. J. Pharm. Sci. 5, 129–137.
Google Scholar
Хаким, К. Р., Тахир, И., Уль Рехман, Р. (2014). Передача сигналов растений: понимание молекулярных перекрестных помех. Индия: Springer 1–355. doi: 10.1007 / 978-81-322-1542-4
CrossRef Полный текст | Google Scholar
Harmer, R. (1990). Связь фаз роста побегов у проростков дуба с развитием стержневого корня, боковых корней и мелких кончиков корней. New Phytol. 115, 23–27. doi: 10.1111 / j.1469-8137.1990.tb00917.x
CrossRef Полный текст | Google Scholar
Hauggaard-Nielsen, H., Gooding, M., Ambus, P., Corre-Hellou, G., Crozat, Y., Dahlmann, C., et al. (2009). Пересечение гороха и ячменя для эффективной симбиотической фиксации N2: получение почвенного азота и использование других питательных веществ в европейских системах органического земледелия. Field Crops Res. 113, 64–71. doi: 10.1016 / j.fcr.2009.04.009
CrossRef Полный текст | Google Scholar
Эрнандес, М., Дюмон, М., Юань, К., Конрад, Р. (2015). Различные популяции бактерий, связанные с корнями и ризосферой риса, включают углерод растительного происхождения. заявл. Environ. Microbiol. 81, 2244–2253. doi: 10.1128 / AEM.03209-14
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Hinsinger, P., Sol, I., Viala, P. (2018). Биодоступность почвенного неорганического фосфора в ризосфере под влиянием химических изменений, вызванных корнями. Растительная почва 237, 173–195.doi: 10.1023 / A: 101335161
CrossRef Полный текст | Google Scholar
Ибрагим, Х., Хатира, А., Пансу, М. (2013). Моделирование функциональной роли микроорганизмов в ежедневном обмене углерода между атмосферой, растениями и почвой. Proc. Environ. Sci. 19, 96–105. doi: 10.1016 / j.proenv.2013.06.011
CrossRef Полный текст | Google Scholar
Икбал, С., Хан, М. Ю., Асгар, Х. Н., Ахтар, М. Дж. (2016). Комбинированное использование солюбилизирующих фосфат бактерий и птичьего помета для увеличения роста и урожайности маша в известковой почве. Soil Environ. 35, 146–154.
Google Scholar
Джамбхулкар П. П., Шарма П., Ядав Р. (2016). Системы доставки для внедрения микробных инокулянтов в поле, в: Микробные инокулянты в устойчивой сельскохозяйственной продуктивности. Springer India Нью-Дели 199–218. doi: 10.1007 / 978-81-322-2644-4_13
CrossRef Полный текст | Google Scholar
Каур, Г., Редди, М. С. (2015). Влияние фосфат-солюбилизирующих бактерий, каменного фосфата и химических удобрений на посевы кукурузы и пшеницы. Педосфера 25, 428–437. doi: 10.1016 / S1002-0160 (15) 30010-2
CrossRef Полный текст | Google Scholar
Khiangte, L., Lalfakzuala, R. (2011). Производство регулятора роста (ИУК) и фосфатазы in vitro бактериями, солюбилизирующими фосфат. Sci. Technol. J. 5, 32–35. doi: 10.22232 / stj.2017.05.01.04
CrossRef Полный текст | Google Scholar
Kondracka, A., Rychter, A. M. (1997). Роль процессов рециклинга P i в процессе фотосинтеза у растений фасоли с дефицитом фосфата. J. Exp. Бот. 48, 1461–1468. doi: 10.1093 / jxb / 48.7.1461
CrossRef Полный текст | Google Scholar
Кумар, А., Сингх, В. К., Трипати, В., Сингх, П. П., Сингх, А. К. (2018). Ризобактерии, способствующие росту растений (PGPR): перспективы в сельском хозяйстве в условиях биотического и абиотического стресса. Урожай Improv. через Microb. Biotechnol. 333–342. doi: 10.1016 / B978-0-444-63987-5.00016-5
CrossRef Полный текст | Google Scholar
Кумар, А. (2016). Фосфатосолюбилизирующие бактерии в сельскохозяйственной биотехнологии: разнообразие, механизм и их роль в росте растений и урожайности. Внутр. J. Adv. Res. 4, 116–124. doi: 10.21474 / IJAR01
CrossRef Полный текст | Google Scholar
Latati, M., Blavet, D., Alkama, N., Laoufi, H. (2014). Посевы вигнового гороха и кукурузы улучшают доступность фосфора в почве и повышают урожайность кукурузы в щелочной почве. Растительная почва 385, 181–191. doi: 10.1007 / s11104-014-2214-6
CrossRef Полный текст | Google Scholar
Latati, M., Bargaz, A., Belarbi, B., Lazali, M., Benlahrech, S., Tellah, S., et al.(2016). Совмещение посевов фасоли с кукурузой улучшает ризобиальную продуктивность, использование ресурсов и урожайность зерна при низкой доступности фосфора. евро. J. Agron. 72, 80–90. doi: 10.1016 / j.eja.2015.09.015
CrossRef Полный текст | Google Scholar
Li, J., Guan, Y., Yuan, L., Hou, J., Wang, C., Liu, F., et al. (2019). Эффекты экзогенной ИУК в регулировании фотосинтетической способности, углеводного обмена и урожайности Zizania latifolia. Sci. Hortic. (Амстердам) 253, 276–285.doi: 10.1016 / j.scienta.2019.04.058
CrossRef Полный текст | Google Scholar
Linu, M. S., Asok, A. K., Thampi, M., Sreekumar, J., Jisha, M. S. (2019). Стимулирующие рост растений признаки местных изолятов pseudomonas aeruginosa, солюбилизирующих фосфат, из ризосферы чили ( capsicumannuum, л). Commun. Почвоведение. Завод анальный. 50, 444–457. doi: 10.1080 / 00103624.2019.1566469
CrossRef Полный текст | Google Scholar
Лю, X., Hegeman, A.Д., Гарднер, Г., Коэн, Дж. Д. (2012). Протокол: высокопроизводительный и количественный анализ ауксина и предшественников ауксина из мельчайших образцов ткани. Plant Methods 8, 31. doi: 10.1186 / 1746-4811-8-31
PubMed Реферат | CrossRef Полный текст | Google Scholar
Лю, X., Jiang, X., He, X., Zhao, W., Cao, Y., Guo, T., et al. (2019). Фосфатсолюбилизирующие pseudomonas sp. штамм p34-l способствует росту пшеницы, заселяя ризосферу пшеницы и улучшая ее корневую систему и фосфорный статус почвы. J. Регулятор роста растений. 38, 1–11. doi: 10.1007 / s00344-019-09935-8
CrossRef Полный текст | Google Scholar
Луис, Дж., Янг, М., Канаширо, С., Джоцис, Т., Рейс, А. (2018). Scientia Horticulturae Silver vase bromeliad: рост растений и минеральное питание при отсутствии макроэлементов. Sci. Hortic. (Амстердам) 234, 318–322. doi: 10.1016 / j.scienta.2018.02.002
CrossRef Полный текст | Google Scholar
Ma, Z., Bielenberg, D. G., Brown, K.М., Линч, Дж. П. (2001). Регулирование плотности корневых волосков за счет наличия фосфора в Arabidopsis thaliana . Среда растительных клеток. 24, 459–467. doi: 10.1111 / pce.12059
CrossRef Полный текст | Google Scholar
Magomya, A. M., Kubmarawa, D., Ndahi, J. A., Yebpella, G. G. (2014). Определение растительных белков с помощью метода Кьельдаля и аминокислотного анализа: сравнительное исследование. Внутр. J. Sci. Technol. Res. 3, 68–72.
Google Scholar
Маджид, А., Аббаси, М. К., Хамид, С., Имран, А., Рахим, Н. (2015). Выделение и характеристика ризобактерий, способствующих росту растений, из ризосферы пшеницы и их влияние на стимуляцию роста растений. Фронт. Microbiol. 6, 198. doi: 10.3389 / fmicb.2015.00198
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мамта Р., Патаниа В., Гулати А., Сингх Б., Бханура Р. К., Тевари Р. (2010). Стимулирующее действие фосфатсолюбилизирующих бактерий на рост растений, содержание стевиозида и ребаудиозида-А в Stevia rebaudiana Bertoni. заявл. Soil Ecol. 46, 222–229. doi: 10.1016 / j.apsoil.2010.08.008
CrossRef Полный текст | Google Scholar
Манзур, М., Аббаси, М. К., Султан, Т. (2017). Выделение солюбилизирующих фосфат бактерий из ризосферы кукурузы и их потенциал для растворения фосфатов – минерализации и стимулирования роста растений. Geomicrobiol. J. 34, 81–95. doi: 10.1080 / 014.2016.1146373
CrossRef Полный текст | Google Scholar
Marathe, R., Патаке, Ю., Шейх, А., Шинде, Б., Гаджбхайе, М. (2017). Влияние ИУК, продуцируемой Pseudomonas aeruginosa 6a (bc4) , на прорастание семян и рост растений Glycin max . J. Exp. Биол. Agric. Sci. 5, 351–358. doi: 10.18006 / 2017.5 (3) .351.358
CrossRef Полный текст | Google Scholar
Midekssa, M. J., Löscher, C. R., Schmitz, R.A., Assefa, F. (2016). Ризобактерии, выделенные из нута ( Cicer aeritinum L.) производственные районы Эфиопии. Afr. J. Biotechnol. 15, 1899–1912. doi: 10.5897 / AJB2015.15172
CrossRef Полный текст | Google Scholar
Мишра, Н., Сундари, С. К. (2013). Нативные PGPM в качестве биоинокулянтов для стимулирования роста растений: ответ на прививку PGPM в основных зерновых и зернобобовых культурах. Внутр. J. Agric. Food Sci. Technol. 4, 1055–1064.
Google Scholar
Мишра, Дж., Сингх, Р., Арора, Н. К. (2017). «Микробы, способствующие росту растений: различные роли в сельском хозяйстве и экологической устойчивости», в Пробиотики и здоровье растений. Изд. Кумар, В., 71–111. doi: 10.1007 / 978-981-10-3473-2
CrossRef Полный текст | Google Scholar
Морган, Дж. А. У., Бендинг, Г. Д., Уайт, П. Дж. (2005). Биологические издержки и преимущества взаимодействия растений и микробов в ризосфере. J. Exp. Бот. 56, 1729–1739. doi: 10.1093 / jxb / eri205
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Оберсон, А., Фризен, Д. К., Рао, И. М., Бюлер, С., Фроссар, Э. (2001). Преобразования фосфора в оксизоле в условиях контрастных систем землепользования: роль микробной биомассы почвы. Растительная почва 2, 197–210. doi: 10.1023 / A: 101330171
CrossRef Полный текст | Google Scholar
Перес, Ю. М., Чарест, К., Далпе, Ю., Сегин, С., Ван, X., Ханизаде, С. (2016). Влияние инокуляции арбускулярными микоризными грибами на отобранные линии яровой пшеницы. Сустейн. Agric. Res. 5, 24. doi: 10.5539 / sar.v5n4p24
CrossRef Полный текст | Google Scholar
Pérez-Miranda, S., Cabirol, N., George-Téllez, R., Zamudio-Rivera, L.S., Fernández, F.Дж. (2007). O-CAS, быстрый и универсальный метод обнаружения сидерофоров. J. Microbiol. Методы 70, 127–131. doi: 10.1016 / j.mimet.2007.03.023
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Перес-Патрисио, С., Кабироль, Н., Камас-Ансуэто, Дж., Санчес-Алегрия, А., Агилар-Гонсалес, А., Гутьеррес-Мичели, Ф. и др. (2018). Оптический метод определения содержания хлорофилла в листьях растений. Sens 18, 650. doi: 10.3390 / s18020650
CrossRef Полный текст | Google Scholar
Pan, X.W., Li, W.-B., Zhang, Q.Y., Li, Y.-H., Liu, M.S. (2008). Оценка характеристик фосфорной эффективности генотипов сои в фосфородефицитных почвах. Сельское хозяйство. Sci. 7, 958–969. doi: 10.1016 / S1671-2927 (08) 60135-2
CrossRef Полный текст | Google Scholar
Панхвар, К. А., Осман, Р., Рахман, З. А., Меон, С. (2011). Вклад фосфатсолюбилизирующих бактерий в биодоступность фосфора и усиление роста аэробного риса. пролет. J. Agric.Res. 9, 810–820. doi: 10.5424 / sjar / 20110903-330-10
CrossRef Полный текст | Google Scholar
Парастеш Ф., Алихани Х. А., Этесами Х. (2019). Биогумус, обогащенный фосфат-солюбилизирующими бактериями, обеспечивает растение достаточным количеством фосфора при последовательном выращивании в условиях известковой почвы. J. Clean Prod. 221, 27–37. doi: 10.1016 / j.jclepro.2019.02.234
CrossRef Полный текст | Google Scholar
Пол, Д., Синха, С. Н. (2017). Выделение и характеристика солюбилизирующей фосфат бактерии Pseudomonas aeruginosa KUPSB12 с антибактериальным потенциалом из реки Ганга, Индия. Ann. Аграр. Sci. 15, 130–136. doi: 10.1016 / j.aasci.2016.10.001
CrossRef Полный текст | Google Scholar
Райя-Гонсалес, Дж., Ортис-Кастро, Р., Руис-Эррера, Л. Ф., Казан, К., Лопес-Бусио, Дж. (2014). Фитохром и время цветения1 / медиатор25 регулируют образование боковых корней посредством передачи сигналов ауксина у арабидопсиса. Plant Physiol. 165, 880–894. doi: 10.1104 / pp.114.239806
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Резахани, Л., Мотешарезаде, Б., Тегерани, М. М., Этесами, Х., Мирсеид Хоссейни, Х. (2019). Фосфат-солюбилизирующие бактерии и кремний синергетически увеличивают поглощение фосфора (P) растением пшеницы ( Triticum aestivum L.), удобренным растворимым или нерастворимым источником фосфора. Ecotoxicol. Environ. Saf. 173, 504–513. doi: 10.1016 / j.ecoenv.2019.02.060
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Розье, К., Герин, Ф., Чарнес, С., Лежандр, Л. (2019). Биозагрузка прорастания кукурузы стимулирующими рост растений ризобактериями Azospirillum lipoferum CRT1. J. Plant Physiol. 237, 111–119. doi: 10.1016 / j.jplph.2019.04.011
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Sarkar, A., Ghosh, P. K., Pramanik, K., Mitra, S., Soren, T., Pandey, S., et al. (2018). Галотолерант Enterobacter sp . проявление активности дезаминазы АСС способствует росту проростков риса в условиях солевого стресса. Res. Microbiol. 169, 20–32. doi: 10.1016 / J.RESMIC.2017.08.005
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Sarsan, S.(2016). Влияние солюбилизирующих фосфат бактерий bacillus psb24 на рост растений томата. Внутр. J. Curr. Microbiol. Прил. Sci. 5, 311–320. doi: 10.20546 / ijcmas.2016.507.033
CrossRef Полный текст | Google Scholar
Шарма, С. Б., Сайед, Р. З., Триведи, М. Х., Гоби, Т. А. (2013). Микробы, солюбилизирующие фосфат: устойчивый подход к управлению дефицитом фосфора в сельскохозяйственных почвах. SpringerPlus 2, 587. doi: 10.1186 / 2193-1801-2-587
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Сингх Р., Сингх В., Сингх П., Ядав Р. А. (2018). Влияние фосфора и ПСБ на показатели урожайности, качество и экономичность летней гринграммы (Vigna radiata L.). Внутр. J. Pharmacogn. Фитохим. Res. 7, 404–408.
Google Scholar
Сонг, Ю. Н., Маршнер, П., Ли, Л., Бао, Х. Г., Сан, Дж. Х., Чжан, Ф. С. (2007). Состав сообществ аммиакокисляющих бактерий в ризосфере промежуточных культур пшеницы ( Triticum aestivum L.), кукурузы ( Zea mays L.) и фасоль бобы ( Vicia faba L.). Biol. Fertil. Почвы 44, 307–314. doi: 10.1007 / s00374-007-0205-y
CrossRef Полный текст | Google Scholar
Сулеман, М., Ид, С. Ю., Расул, М., Яхья, М., Атта, М., Мирза, М. С. (2018). Фосфатосолюбилизирующие бактерии с геном глюкозодегидрогеназы для поглощения фосфора и положительного воздействия на пшеницу. PloS One 13, 1–28. doi: 10.1371 / journal.pone.0204408
CrossRef Полный текст | Google Scholar
Sun, Y.М., Чжан, Н. Н., Ван, Э. Т., Юань, Х. Л., Ян, Дж. С., Чен, В. X. (2009). Влияние посевов и посевов плюс посев ризобий на микробную активность и состав сообществ в ризосфере люцерны ( Medicago sativa L.) и сибирской дикой ржи ( Elymus sibiricus L.). FEMS Microbiol. Ecol. 70, 218–226. doi: 10.1111 / j.1574-6941.2009.00752.x
CrossRef Полный текст | Google Scholar
Тахир, М., Халид, У., Иджаз, М., Мустафа, Г., Карим Ф., Наим М. и др. (2018). Комбинированное применение биоорганических фосфатных и фосфорсолюбилизирующих бактерий (штамм Bacillus MWT 14 ) улучшает урожайность мягкой пшеницы при низком внесении удобрений в засушливых условиях. Braz. J. Microbiol. 49, 1–10. doi: 10.1016 / j.bjm.2017.11.005
CrossRef Полный текст | Google Scholar
Тан, А., Харуна, А. О., Мухамад, Н., Маджид, А. (2018). Возможные свойства PGPR целлюлолитических, азотфиксирующих и фосфатсолюбилизирующих бактерий восстановленной почвы тропического леса. BioRxiv 8, 1–60. doi: 10.1101 / 351916
CrossRef Полный текст | Google Scholar
Тарик, А., Сабир, М., Фарук, М., Максуд, М.А., Ахмад, Х.Р., Варрайч, Э.А. (2014). «Дефицит фосфора в растениях: ответы, адаптивные механизмы и передача сигналов», в Plant Signaling: Understanding the Molecular Crosstalk , 133–148. doi: 10.1007 / 978-81-322-1542-4
CrossRef Полный текст | Google Scholar
Vance, C.P., Vance, C.P., Uhde-stone, C., Аллан, Д. Л. (2003). Приобретение и использование фосфора: критическая адаптация растений для обеспечения невозобновляемого ресурса. New Phytol. 157, 423–447. doi: 10.1046 / j.1469-8137.2003.00695.x
CrossRef Полный текст | Google Scholar
Vandamme, E., Wissuwa, M., Rose, T., Ahouanton, K., Saito, K. (2016). Стратегическое внесение фосфора (P) в грядку питомника увеличивает рост рассады и урожай пересаженного риса при низком содержании фосфора. F. Crop Res. 186, 10–17.doi: 10.1016 / j.fcr.2015.11.003
CrossRef Полный текст | Google Scholar
Ван, Дж., Чу, Г. (2015). Форма и стратегия применения фосфорных удобрений влияют на подвижность и трансформацию фосфора в карбонатной почве с капельным орошением. J. Plant Nutr. Почвенный Sc. 178, 914–922. doi: 10.1002 / jpln.201500339
CrossRef Полный текст | Google Scholar
Ван, Б., Мэй, К., Зайлер, Дж. Р. (2015). Стимуляция раннего роста и изменения физиологии на уровне листьев у инокулированного штамма Burkholderia phytofirmans PsJN проса. Plant Physiol. Bioch. 86, 16–23. doi: 10.1016 / j.plaphy.2014.11.008
CrossRef Полный текст | Google Scholar
Wang, F., Shi, N., Jiang, R., Zhang, F., Feng, G. (2016). Исследование стабильных изотопов in situ фосфат-солюбилизирующих бактерий в гифосфере. J. Exp. Бот. 67, 1689–1701. doi: 10.1093 / jxb / erv561
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Wang, Q., Ye, J., Wu, Y., Luo, S., Chen, B., Ma, L., et al. (2019).Стимулирование развития корней и поглощения Zn Sedum alfredii было достигнуто эндофитной бактерией Sasm05 . Ecotoxicol. Environ. Saf. 172, 97–104. doi: 10.1016 / j.ecoenv.2019.01.009
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Вани, П., Заиди, А., Хан, А., Хан, М. С. (2005). Влияние фората на солюбилизацию фосфата и потенциал высвобождения индолуксусной кислоты ризосферными микроорганизмами. Ann. Plant Prot. 13, 139–144.
Google Scholar
Цзэн, Ф., Шабала, Л., Чжоу, М., Чжан, Г., Шабала, С. (2013). Реакции ячменя на комбинированный стресс от заболачивания и засоления: разделение эффектов кислородного голодания и элементарной токсичности. Фронт. Plant Sci. 4, 313. doi: 10.3389 / fpls.2013.00313
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Zheng, L.Q., Narsai, R., Wu, J.J., Giraud, E., He, F., Cheng, L.J. и др. (2009). Физиологический и транскриптомный анализ взаимодействия железа и фосфора в проростках риса. Plant Physiol. 151, 262–274. doi: 10.1104 / pp.109.141051
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Фосфатосолюбилизирующие бактерии сводят на нет антагонистический эффект кальцификации почвы на биодоступность фосфора в щелочных почвах
Чен, З., Ма, С. и Лю, Л.Л. Исследования активности штамма фосфобактерий по солюбилизации фосфора изолирован из почвы каштанового типа в Китае. Bioresour Technol. 99 , 6702–6707 (2008).
CAS Статья PubMed Google ученый
Абди Д., Кейд-Менун Б. Дж., Зиади Н. и Родитель Л. Э. Долгосрочное влияние методов обработки почвы и внесения фосфорных удобрений на формы фосфора в почве по данным спектроскопии ядерного магнитного резонанса. J.Environ. Qual. 43 , 1431–1441 (2014).
Артикул PubMed Google ученый
Halajnia, A., Haghnia, G.H., Fotovat, A. & Khorasani, R. Фракции фосфора в известняковых почвах с внесением удобрений P и навоза крупного рогатого скота. Геодерма. 150 , 209–213 (2009).
ADS CAS Статья Google ученый
Байер, К., Мартин-Нето, Л., Мельничук, Дж. И Сангой, Л. Изменения фракций органического вещества почвы в субтропических системах земледелия с нулевой обработкой почвы. Общество почвоведов Америка Дж. 65 , 1473–1478 (2001).
ADS CAS Статья Google ученый
Ядав, Х., Фатима, Р., Шарма, А. и Матур, С. Повышение применимости каменного фосфата в щелочных почвах с помощью органического компоста. Appl Soil Ecol. 113 , 80–85 (2017).
Артикул Google ученый
Rehim, A. et al. .Ленточное внесение фосфора с фермерским навозом улучшает эффективность использования фосфора, урожайность и чистую отдачу пшеницы на супесчаной глинистой почве. Turk J Agric For. 40 , 319–326 (2016).
Артикул Google ученый
Шариф М., Хаттак Р. А. и Сарир М. С. Влияние различных уровней гуминовой кислоты, полученной из лигнитного угля, на рост растений кукурузы. Commun. В почв. и заводить анал.СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 33 , 1920–1924 (2002).
Google ученый
Лопес-Бусио, Дж., Де ла Вега, О. М., Гевара-Гарсиа, А. и Эррера-Эстрелла, Л. Повышенное поглощение фосфора трансгенными растениями табака, которые продуцируют избыточное количество цитрата. Нац. Биотех. 18 , 450–453 (2000).
Артикул Google ученый
Сато, С., Соломон, Д., Hyl, C., Ketterings, Q. M. & Lehmann, J. Видообразование фосфора в навозе и почвах с добавками навоза с использованием спектроскопии XANES. Environ. Sci. Technol. 39 , 7485–7491 (2005).
ADS CAS Статья PubMed Google ученый
Алам М. и Ладха Дж. К. Оптимизация фосфорных удобрений в системе интенсивного выращивания овощей и риса. Biol. Fert. Почвы. 40 , 277 283 (2004).
Артикул Google ученый
Weyers, E. et al. . Phosphorus Speciat ion в известковых почвах после ежегодных поправок к молочному навозу. Почвоведение. Soc Am. 80 , 1531–1542 (2016).
CAS Статья Google ученый
Редди М. С., Кумар С., Бабита К. и Редди М. С. Биосолюбилизация плохо растворимых фосфатов горных пород с помощью Aspergillus tubingensis и Aspergillus niger. Biresource Technol. 84 , 187–189 (2002).
CAS Статья Google ученый
Шарма, С. Б., Сайед, Р. З., Триведи, М. Х. и Гоби, Т. А. Микробы, солюбилизирующие фосфор: устойчивый подход к управлению дефицитом фосфора в сельскохозяйственных почвах. Springer Plus. 2 , 587 (2013).
Артикул PubMed PubMed Central Google ученый
Чен, Ю. П. и др. . Солюбилизирующие фосфат бактерии из субтропических почв и их способность растворять трикальций. заявл. Soil Ecol. 34 , 33–41 (2006).
Артикул Google ученый
Родригес, Х. и Фрага, Р. Фосфатосолюбилизирующие бактерии и их роль в стимулировании роста растений. Biotechnol. Adv. 17 , 319–339 (1999).
CAS Статья PubMed Google ученый
Сяо, Ю., Ван, X., Чен, В. и Хуанг, К. Выделение и идентификация трех солюбилизирующих калий бактерий из ризосферной почвы рапса и их влияние на райграс. Геомикробиологический журнал . 1–8 (2017).
Сугихара, С., Фунакава, С., Киласара, М. и Косаки, Т. Динамика азота микробной биомассы в зависимости от поглощения азота растениями в период роста сельскохозяйственных культур в засушливых тропических пахотных землях Танзании. Почвоведение. Завод орех. 56 , 105–114 (2010).
CAS Статья Google ученый
Раджан, С.С., Уоткинсон, Дж. Х. и Синклер, А. Г. Фосфатные породы для непосредственного внесения в почву. Adv. Агрон. 57 , 77–159 (1996).
CAS Статья Google ученый
Зайда А., Хан, М. С. и Амил, М. Д. Интерактивное влияние ризотрофных микроорганизмов на урожайность и потребление питательных веществ нута (Cicer arietinum L.). евро. J. Agron. 19 , 15–21 (2003).
Артикул Google ученый
Лугтенберг, Б., Чин-А-Вунг, Т. и Блумберг, Г. Взаимодействие микробов и растений: принципы и механизмы. Антони ван Левенгук. 81 , 373–383 (2002).
CAS Статья PubMed Google ученый
Хаса, Ю. П., Баббал и Адивития.Микробы как агенты биоконтроля. В Probiotics andPlant Health (стр. 507-552). Springer Singapore (2017).
Айер, Б. и Раджкумар, С. Специфичность хозяина и стимуляция роста растений бактериальными эндофитами. Curr Res Microbiol Biotechnol. 5 , 1018–1030 (2017).
Google ученый
Аль-Энази, А. А. Р., Аль-Уд, С. С., Аль-Барака, Ф. Н. и Усман, А. Р. Роль микробной инокуляции и промышленного побочного фосфогипса в росте и усвоении питательных веществ кукурузы (Zea mays L.) выращивают на известковой почве. J. Sci. Еда. Agri. 97 (11), 3665–3674 (2017).
Артикул Google ученый
Ким К. Ю., Джордан Д. и Макдональд Г. А. Солюбилизация гидроксиапатита Enterobacter agglomerans и клонирование Escherichia coli в культуральной среде. Biol. Fert. Почвы. 24 , 347–352 (1997).
CAS Статья Google ученый
Рашид, М., Самина, К., Наджма, А., Садия, А. и Фарук, Л. Производство органических кислот и солюбилизация фосфата солюбилизирующими фосфат микроорганизмами в условиях in vitro . пак. J. Biol. Sci. 7 , 187–196 (2004).
Артикул Google ученый
Бхаттарай К. П. и Мандал Т. Н. Взаимосвязь между микробной биомассой почвы и тонкими корнями в Тарае и Хилл Сал (Shorea robusta Gaertn.) Леса Восточного Непала. Внутр. J. Ecol. Environ Sci. 42 , 19–28 (2017).
Google ученый
Гулати А. и др. . Производство органических кислот и стимуляция роста растений в зависимости от солюбилизации фосфата штаммом Acinetobacter rhizosphaerae BIHB 723, выделенным из холодных пустынь трансгималаев. Arch. Microbiol. 192 , 975–983 (2010).
CAS Статья PubMed Google ученый
Oliveira, C.A. et al. . Солюбилизирующие фосфат микроорганизмы, выделенные из ризосферы кукурузы, культивируемой в оксизоле бразильского биома Серрадо. Soil Biol Biochem. 41 , 1782–1787 (2009).
CAS Статья Google ученый
Алори, Э. Т., Глик, Б. Р. и Бабалола, О. О. Солюбилизация микробного фосфора и его потенциал для использования в устойчивом сельском хозяйстве. Передняя . Микробиол . 8, https://doi.org/10.3389/fmicb.2017.00971 (2017).
Джонс, Д., Смит, Б. Ф. Л., Уилсон, М. Дж. И Гудман, Б. А. Фосфат-солюбилизирующие грибы в шотландской высокогорной почве. Mycol. Res. 95 , 1090–1903 (1991).
Артикул Google ученый
Стивенсон Ф. Дж. Циклы почвы: углерод, азот, фосфор, сера, микроэлементы. Джон Уайли и сыновья, Нью-Йорк (2005).
Deubel, A., Gransee & Merbach, W. Трансформация органических ризопланов бактериями ризопланы и ее влияние на доступность третичного фосфата кальция. J. Plant Nutr. Почвоведение. 163 , 387–392 (2000).
CAS Статья Google ученый
Deubel, A. & Merbach, W. Влияние микроорганизмов на биодоступность фосфора в почвах. В: Бускот, Ф.и А. Варма (ред.). Микроорганизмы в почвах: роль в генезисе и функциях , стр: 177–91 Springer-Verlag, Берлин-Гейдельберг, Германия (2005).
Хан, М. и Шариф. M Повышение растворимости фосфора из каменного фосфата путем компостирования с подстилкой для домашней птицы. Sarhad J. Agric. 28 , 415–420 (2012).
Google ученый
Mehta, P., Walia, A., Kulshrestha, S., Chauhan, A. & Shirkot, C.K. Эффективность стимулирующего рост растений P-солюбилизатора Bacillus circus CB7 для усиления роста томатов в чистых домашних условиях. J. Basic Microb. 53 , 1–12 (2014).
Google ученый
Хан, А. А., Джилани, Г., Ахтер, М. С., Накви, С. М. С. и Рашид, М. Бактерии, солюбилизирующие фосфор; Возникновение, механизмы и их роль в растениеводстве. J. Agric. Биол. Sci. 1 , 48–58 (2009).
Google ученый
Begum, M., Narayanasamy, G. & Biswas, D. R. Способность фосфора обеспечивать фосфатные породы под влиянием уплотнения водорастворимыми P удобрениями, Nutr. Цикл. Агроэкосис. 68 , 73–84 (2004).
CAS Статья Google ученый
Mohammadi, K., Ghalavand, A., Aghaalikhani, M., Heidari, G. R. & Sohrabi, Y., знакомство с системой устойчивого плодородия почвы для нута (Cicer arietinum L.). Afr J. Biotech. 10 , 6011–6020 (2011).
Google ученый
Салим, М., Аршад, М. и Ясин, М. Эффективность различных подходов к использованию каменного фосфата в качестве потенциального источника доступного для растений фосфора для устойчивого производства пшеницы. Внутр. J. Agric. Биол. 15 , 223–230 (2013).
CAS Google ученый
Хао, Х., Чо, К. М., Рач, Дж. Дж. И Чанг, К. Химическое замедление диффузии фосфатов в кислой почве под воздействием известкования. Nutr. Цикл. Агроэкосис. 64 , 213–224 (2002).
CAS Статья Google ученый
Линдси, В.Л. П. Л. Г. В и Чиен, С. Х. Фосфатные минералы. Фосфатные минералы. Гл. 22, стр. 1089-1130. В: J.B. Dixon and S.B. Сорняк (ред.), Минералы в почвенной среде, 2-е издание (1989).
Барроу, Н. Дж. Влияние pH на поглощение фосфатов из почвы. Растение и почва. 410 , 401–410 (2017).
CAS Статья Google ученый
Salvagiotti, F. et al. . S-стехиометрия зерна и физиологические признаки, связанные с урожаем зерна кукурузы под влиянием фосфорного и серного питания. Field Crops Res. 203 , 128–138 (2017).
Артикул Google ученый
Шен, К., Хедли, М., Кэмпс Арбестейн, М. и Киршбаум, М. У. Ф. Может ли biochar увеличить биодоступность фосфора? J. Почвоведение. Завод Нутр. 16 , 268–286 (2016).
Google ученый
Curtin, D. & Syers, J. K.Вызванные известью изменения показателей доступности почвенного фосфата. Почвоведение. Soc. Являюсь. J. 65 , 147–152, https://doi.org/10.2136/sssaj2001.651147x (2001).
ADS CAS Статья Google ученый
Альварес Р., Эванс Л. А., Милхэм П. Дж. И Уилсон М. А. Влияние гуминовых веществ на осаждение фосфата кальция. Геодерма. 118 , 245–260 (2004).
ADS CAS Статья Google ученый
Дельгадо, А., Мадрид, А., Кассем, С., Андреу, Л. и дель Кампильо, М. С. Извлечение фосфорных удобрений из известковых почв с добавлением гуминовых и фульвокислот. Растительная почва. 245 , 277–286 (2002).
CAS Статья Google ученый
Гроссл П. и Инскип У. П. Осаждение дигидрата дикальцийфосфата в присутствии органических кислот. Почвоведение. Soc. Являюсь. J. 55 , 670–675 (1991).
ADS CAS Статья Google ученый
Сатьяпракаш, М., Никита, Т., Редди, Э. У. Б., Садхана, Б. и Вани, С. С. Бактерии, растворяющие фосфор и фосфат, и их роль в питании растений. Внутр. J. Curr. Microbiol. Приложение. Sci. 6 , 2133–2144 (2017).
Артикул Google ученый
Гольдштейн, А. Х. Бактериальная солюбилизация минеральных фосфатов: историческая перспектива и перспективы на будущее. Am J Altern Agri. 1 , 51–7 (1986).
Артикул Google ученый
Kim, K. Y., Jordan, D. & Krishnan, H. B. Rahnella aqualitis, бактерия, выделенная из ризосферы сои, может солюбилизировать гидроксиапатит. FEMS Microbiol. Lett. 153 , 273–277 (1997).
CAS Статья Google ученый
Джонс, Д.Л. Органические кислоты в ризосфере – критический обзор. Почва растений 205 , 25–44 (1998).
CAS Статья Google ученый
Maliha, R., Samina, K., Najma, A., Sadia, A. & Farooq, L. Производство органических кислот и солюбилизация фосфата солюбилизирующими фосфат микроорганизмами в условиях in vitro . Pak J. Biol. Sci. 7 , 187–196 (2004).
Артикул Google ученый
Хан, М. С., Заиди, А. и Вани, П. А. Роль солюбилизирующих фосфат микроорганизмов в устойчивом сельском хозяйстве – обзор. Агрон. Поддерживать. Dev. 26 , 1–15 (2006).
Артикул Google ученый
Taktek, S., St-Arnaud, M., Piché, Y., Fortin, J. A. & Antoun, H. Солюбилизация магматических фосфатных пород с помощью образующих биопленку микоризобактерий и гифобактерий, связанных с Rhizoglomus irregulare DAOM 197198. Микориза. 27 , 13–22 (2017).
CAS Статья PubMed Google ученый
Wu, S.C., Cao, Z.H., Li, Z.G., Cheung, K.C. & Wong, M.H. Влияние биоудобрения, содержащего N-фиксатор, солюбилизаторы P и K и AM-грибы, на рост кукурузы: тепличное испытание. Геодерма. 125 , 155–166 (2005).
ADS Статья Google ученый
Кумар С., Баудд К., Барман С. и Сингх Р. П. Внесение микробных биоудобрений и органических веществ снижает потребность в мочевине и DAP за счет повышения доступности питательных веществ и урожайности пшеницы (Triticum aestivum L.). Ecol. Англ. 71 , 432–437 (2014).
Артикул Google ученый
Махди, С. С., Хассан, Г. И., Хуссейн, А. и Расул, Ф. Проблема доступности фосфора – его фиксация и роль солюбилизирующих фосфат бактерий в солюбилизации фосфата. Res. J. Agric. Sci. 2 , 174–179 (2011).
Google ученый
Пахари А. и Мишра Б. Б. Характеристика ризобактерий, продуцирующих сидерофор, и его влияние на показатели роста различных овощей. Внутр. J. Curr. Microbiol. Приложение. Sci. 6 (5), 1398–1405 (2017).
Артикул Google ученый
Хамида, Б., Харини, Г., Рупела, О. П. и Вани, С. П. Гопал Редди. Стимуляция роста кукурузы фосфатосолюбилизирующими бактериями, выделенными из компостов и макрофауны. Microbiol. Res. 163 , 234–242 (2008).
CAS Статья PubMed Google ученый
Нарди, Серенелла, Диего Пиццегелло и Андреа Эртани. «Гормоноподобная активность органического вещества почвы». Appl Soil Ecol ., DOI: https: //doi.org/10.1016/j.apsoil.2017.04.020 (2017).
Байс, Х. П., Лойола-Варгас, В. М., Флорес, Х. Э. и Виванко, Дж. М. Корневоспецифический метаболизм: биология и биохимия подземных органов. In Vitro Cell Dev. Biol. Растение. 37 , 30–741 (2001).
Google ученый
Six, J. et al. . Источники и состав почвенного органического вещества, фракции между почвенными агрегатами и внутри них. Eur J. Soil Sci. 52 , 607–618 (2001).
CAS Статья Google ученый
Gyaneshwar, P. et al. . Участие индуцируемой фосфатным голоданием глюкозодегидрогеназы в солюбилизации фосфатов в почве с помощью Enterobacter asburiae. FEMS. Microbiol. Lett. 171 , 223–229 (1999).
CAS Статья Google ученый
Криг, Н. Р. и Холт, Дж. Г. (ред.). Руководство Гергея по систематической бактериологии. 9-е издание. Vol. 1. Уильямс и Уилкинс, Балтимор, Лондон. 984pp (1984).
Холт, Дж. Г., Криг, Н. Р., Снит, П. Х. А., Стейли, Дж. Т. и Уильямс, С. Т. (ред.). Руководство Берджи по детерминантной бактериологии. 9-е издание. Уильямс и Уилкин, Балтимор. MD. 787pp (1994).
Гордон Р. Э., Хейнс В. К. и Панг К. Н. Род Bacillu s.Справочник по сельскому хозяйству. № 427. Министерство сельского хозяйства США, Вашингтон, округ Колумбия, 283 стр. (1973).
Bashan, Y., Kamnev, A. A. & de-Bashan, L.E. Трикальцийфосфат не подходит в качестве универсального фактора отбора для выделения и тестирования фосфат-солюбилизирующих бактерий, усиливающих рост растений: предложение по альтернативной процедуре. Биол . Фертиль . Почвы . 1–15 (2013).
Кумар Р. и Шастри Б.Роль фосфатсолюбилизирующих микроорганизмов в устойчивом развитии сельского хозяйства. В Агроэкологическая устойчивость (стр. 271–303). Издательство Springer International (2017).
Томас, Г. У. pH почвы и кислотность почвы. В Спаркс Д. и др. . Методы анализа почв: Часть 3, Химические методы. Мэдисон: Почвоведение. Soc Am . 475–490 (1996).
Роудс, Дж. Д. Соленость: электрическая проводимость и общее количество растворенных твердых веществ.В: Sparks D, Page A, Helmke P, et al . Методы анализа почв: Часть 3, Химические методы. Мэдисон : Soil Sci Soc Am J . 417–435 (1996).
Малвани Р. Л. 1996. Неорганические формы азота. В Д.Л. Спарк, (ред.) Метод анализа почвы, часть 3: Химические методы серии книг SSSA № 6 SSSA, I NC ., ASA, I NC ., Мэдисон, Висонсин, США. 1123–1184pp (1996).
Райан, Дж., Эстефан, Г.И Рашид, А. Руководство лаборатории анализа почвы и растений (2-е изд.). Совместно опубликовано Международным центром сельскохозяйственных исследований в засушливых регионах (ICARDA) и Национальным центром сельскохозяйственных исследований (NARC). Доступно в ИКАРДА, Алеппо, Сирия. 172 стр. (2001).
Олсен, С. Р., Коул, К. В., Ватанабе, Ф. С. и Дин, Л. А. Оценка доступного фосфора в почвах путем экстракции бикарбонатом натрия. Вашингтон, округ Колумбия: Типография правительства США (1954).
Loeppert, R. H & Suarez, D. L. Карбонаты и гипс. В Спаркс, Д., Пейдж, А., Хельмке, П. и др. . Методы анализа почв: Часть 3, Химические методы. Мэдисон : Soil Sci Soc Am J . 437–474 (1996).
Джи, Г. У. и Баудер, Дж. У. Анализ размера частиц. п. 383–411. В Клют, А. (ред.) Методы анализа почвы. Часть 1. 2-е изд. Агрон. Monogr. 9. ASA и SSSA, Мэдисон, Висконсин (1986).
Нельсон Д. У. и Соммерс Л. Э. Общий углерод, органический углерод и органические вещества. В: Методы анализа почвы, часть 2, 2-е изд., Пейдж, А. Л. и др. . Эд. Агрономия. 9: 961–1010. Soc. компании Agron., Inc., Мэдисон, Висконсин (1996).
Nautiyal, C. S. Эффективная микробиологическая питательная среда для скрининга микроорганизмов, растворяющих фосфаты. Письма по микробиологии FEMS. 170 , 265–270 (1999).
CAS Статья PubMed Google ученый
Александр, Д. Б. и Зуберер, Д. А. Использование реагентов хром-азурол S для оценки продукции сидерофоров ризосферными бактериями. Biol. Fertil. Почвы. 12 , 39–45 (1991).
CAS Статья Google ученый
Винсет, Дж. М. А. Руководство по практическому изучению корневых клубеньковых бактерий. IBPH и книга № 15. Научная публикация Блэквелла, Оксфорд (1970).
Эйвази, Ф.& Табатабай, М. А. Фосфатазы в почвах. Soil Biol. Biochem. 9 , 167–172 (1977).
CAS Статья Google ученый
Пиковская Р.И. Мобилизация фосфатов в почве в связи с жизнедеятельностью некоторых видов микробов. Микробиология. 17 , 362–370 (1948).
CAS Google ученый
Стил Р. Д. и Тор Дж. Х. Принципы и процедуры статистики. Макгроу-Хилл, Торонто, 481 стр (1960).
Влияние бактериальных инокулянтов, солюбилизирующих ризобий и фосфат, на симбиотические признаки, леггемоглобин клубеньков и урожай генотипов нута
В ходе сезона раби 2004-05 годов был проведен полевой эксперимент, чтобы выяснить влияние ризобия и инокулянты фосфорсолюбилизирующих бактерий (PSB) на симбиотические признаки, леггемоглобин клубеньков и урожай пяти элитных генотипов нута.Среди генотипов нута IG-593 показал лучшие результаты в отношении симбиотических параметров, включая количество клубеньков, свежий вес клубеньков, сухой вес клубеньков, сухой вес побегов, характеристики урожайности и урожайность. Содержание леггемоглобина (2,55 мг г -1 свежих клубеньков) также было выше при IG-593. Среди микробных инокулянтов наиболее эффективным оказался Rhizobium + PSB с точки зрения количества клубеньков (27,66 клубеньков, растение -1 ), массы свежих клубеньков (144,90 мг растения -1 ), сухой массы клубеньков (74.30 мг растения -1 ), сухой массы побегов (11,76 г растения -1 ) и содержания леггемоглобина (2,29 мг г -1 свежих клубеньков), а также продемонстрировали свой положительный эффект в повышении всех параметров, определяющих урожай. , урожай зерна и соломы.
1. Введение
Зернобобовые – вторая по важности группа сельскохозяйственных культур после зерновых. Развивающиеся страны вносят около 74% в мировое производство зернобобовых, а оставшаяся часть приходится на развитые страны. Индия, Китай, Бразилия, Канада, Мьянма и Австралия являются основными странами-производителями зернобобовых с относительной долей 25%, 10%, 5%, 5% и 4% соответственно.В 2009 году мировое производство зернобобовых составило 61,5 млн тонн с площади 70,6 млн га при средней урожайности 871 кг / га. На сухие бобы приходилось около 32% мирового производства зернобобовых, за ними следуют сухой горох (17%), нут (15,9%), фасоль (7,5%), чечевица (5,7%), вигна (6%) и голубиный горох (4,0%). . Индия является крупнейшим производителем и потребителем зернобобовых в мире, на ее долю приходится около 25–28% от общего мирового производства. Около 75% мирового нута ( Cicer arietinum L.) область приходится на Индию [1]. Нут – одна из основных послеводожденных сезонных культур в Мадхья-Прадеше, занимающая 3,09 млн га при урожайности 3,30 млн тонн и урожайности 1071 кг га −1 [2]. Низкая урожайность нута в этом регионе в основном связана с несбалансированным внесением питательных веществ и использованием традиционных сортов. В таких ситуациях использование Rhizobium и фосфатсолюбилизирующих бактерий (PSB) показало преимущество в повышении продуктивности нута [3, 4].Микробные инокулянты являются экономичными, экологичными и возобновляемыми источниками питательных веществ для растений [5]. Rhizobium и PSB имеют большое значение из-за их жизненно важной роли в -фиксации N 2 и P-солюбилизации. Введение эффективных штаммов P-солюбилизирующих видов Bacillus megaterium , биовара phosphaticum , Bacillus polymyxa , Pseudomonas striata , Aspergillus awamori и Penicizillium, было зарегистрировано в посевах rigitalium и Penicizillium. для увеличения доступности фосфора в почве [6].Поскольку информация о реакции элитных генотипов нута на инокуляцию Rhizobium и фосфатосолюбилизирующими бактериальными инокулянтами в такой ситуации скудна, был разработан эксперимент для оценки продуктивности генотипов нута в комбинациях с микробными инокулянтами в регионе Мальва . Мадхья-Прадеш.
2. Материалы и методы
2.1. Экспериментальная площадка
Полевой эксперимент проводился на экспериментальной ферме сельскохозяйственного колледжа, Индор, Мадхья-Прадеш (22 ∘ 43 ′ северной широты, 75 ∘ 56 ′ восточной долготы и 555.7 м над средним уровнем моря). Почва опытного участка относится к сароловой серии, входящей в состав тонкого монтмориллонитового семейства вертик-устохрептов и вертик-хромустеров. Характеристики почвы экспериментального участка перед началом исследования были проанализированы и представлены в таблице 1.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||