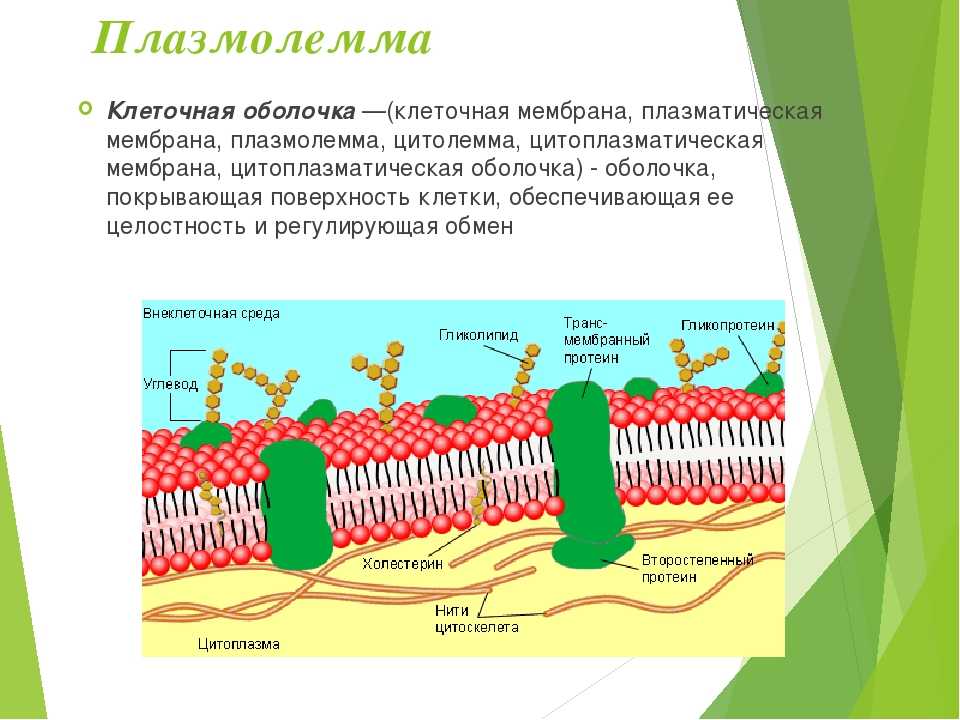
Клеточная мембрана – наружная структура, отличие от клеточной стенки, жидкостно-мозаичная модель
4.7
Средняя оценка: 4.7
Всего получено оценок: 1496.
4.7
Средняя оценка: 4.7
Всего получено оценок: 1496.
Клеточная мембрана – это плоскостная структура, из которой построена клетка. Она присутствует у всех организмов. Её уникальные свойства обеспечивают жизнедеятельность клеток.
Виды мембран
Можно выделить три вида клеточных мембран:
- наружная;
- ядерная;
- мембраны органоидов.
Наружная цитоплазматическая мембрана создаёт границы клетки. Её не надо путать с клеточной стенкой или оболочкой, имеющейся у растений, грибов и бактерий.
Отличие клеточной стенки от клеточной мембраны в значительно большей толщине и преобладании защитной функции над обменной. Мембрана располагается под клеточной стенкой.
Ядерная мембрана отделяет от цитоплазмы содержимое ядра.
Среди органоидов клетки есть такие, форма которых образована одной или двумя мембранами:
- митохондрии;
- пластиды;
- вакуоли;
- комплекс Гольджи;
- лизосомы;
- эндоплазматическая сеть (ЭПС).
Строение мембраны
По современным представлениям структура клеточной мембраны описывается с помощью жидкостномозаичной модели. Основу мембраны составляет билипидный слой – два уровня молекул липидов, образующих плоскость. С обеих сторон на билипидном слое расположены молекулы белков. Некоторые белки погружены в билипидный слой, некоторые проходят через него.
Рис. 1. Клеточная мембрана.Животные клетки на поверхности мембраны имеют комплекс углеводов. При изучении клетки под микроскопом отмечено, что мембрана находится в постоянном движении и неоднородна по строению.
Мембрана является мозаикой и в морфологическом, и в функциональном смысле, т. к. её различные участки содержат различные вещества и имеют разные физиологические свойства.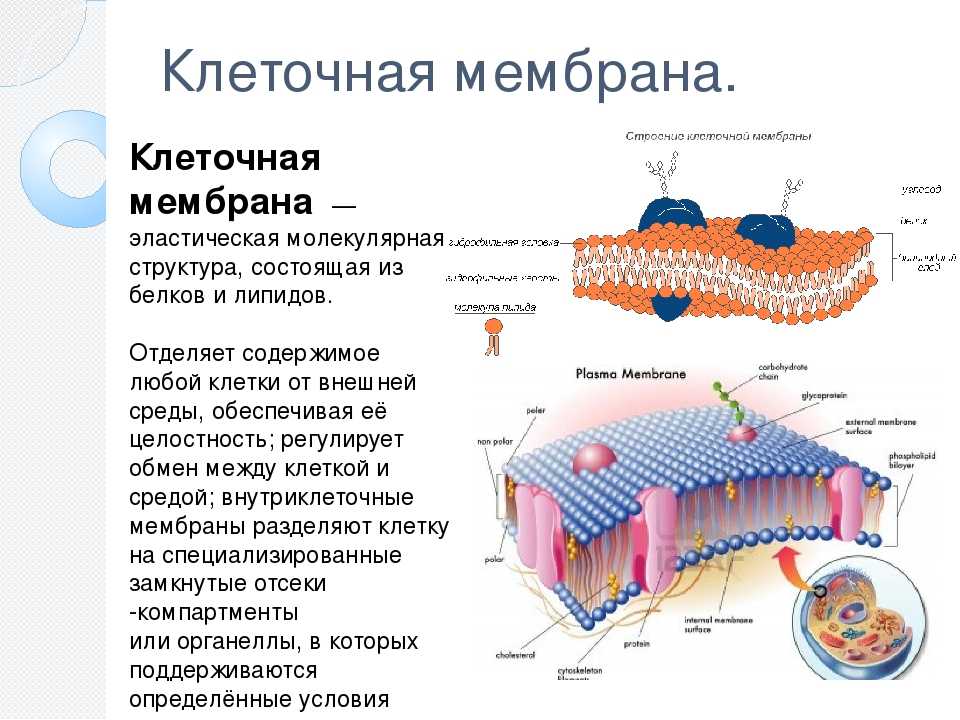
Свойства и функции
Любая пограничная структура осуществляет защитные и обменные функции. Это касается и всех видов мембран.
Осуществлению данных функций способствуют такие свойства, как:
- пластичность;
- высокая способность к восстановлению;
- полупроницаемость.
Свойство полупроницаемости заключается в том, что одни вещества не пропускаются мембраной, а другие пропускаются свободно. Так осуществляется контролирующая функция мембраны.
Также наружная мембрана обеспечивает связь между клетками за счёт многочисленных выростов и выделения клеящего вещества, заполняющего межклеточное пространство.
Транспорт веществ через мембрану
Поступление веществ через наружную мембрану идёт следующими путями:
- через поры с помощью ферментов;
- через мембрану непосредственно;
- пиноцитозом;
- фагоцитозом.
Первыми двумя способами транспортируются ионы и мелкие молекулы. Крупные молекулы поступают в клетку путём пиноцитоза (в жидком состоянии) и фагоцитоза (в твёрдом виде).
Мембрана обхватывает пищевую частицу и замыкает её в пищеварительную вакуоль.
Вода и ионы проходят в клетку без затрат энергии, пассивным транспортом. Крупные молекулы перемещаются активным транспортом, с затратой энергетических ресурсов.
Внутриклеточный транспорт
От 30 % до 50 % объёма клетки занимает эндоплазматическая сеть. Это своеобразная система полостей и каналов, связывающая все части клетки и обеспечивающая упорядоченную внутриклеточную транспортировку веществ.
Рис. 3. Рисунок ЭПС.Таким образом, в ЭПС сосредоточена значительная масса клеточных мембран.
Что мы узнали?
Мы выяснили что такое клеточная мембрана в биологии. Это структура, на основе которой построены все живые клетки. Её значение в клетке заключается в: отграничении пространства органоидов, ядра и клетки в целом, обеспечении избирательного поступления веществ в клетку и ядро. В состав мембраны входят молекулы липидов и белков.
Тест по теме
Доска почёта
Чтобы попасть сюда – пройдите тест.
Александр Котков
10/10
Василий Курамшин
10/10
Полина Ратова
7/10
Оценка доклада
4.7
Средняя оценка: 4.7
Всего получено оценок: 1496.
А какая ваша оценка?
21. Наружная мембрана
19.
Внутренние мембраны клетки, несмотря
на различия в морфологическом и
функциональном отношениях, представляют
собой единое целое, поскольку способны
переходить друг в друга в процессе
функционирования клетки как будет
показано ниже. К вакуолярной системе
относятся МЕМБРАНЫ ЭНДОПЛАЗМАТИЧЕСКОГО
РЕТИКУЛУМА, АППАРАТА ГОЛЬДЖИ, МИКРОТЕЛЬЦА,
ЛИЗОСОМЫ и ВАКУОЛИ РАСТИТЕЛЬНОЙ
КЛЕТКИ.
Мембраны
эндоплазматического ретикулума являются
крайне сложным и разветвленным
продолжением внешней мембраны
двухмембранной ядерной оболочки (рис.
74). Мембраны эндоплазматического
ретикулума (ЭПР) в типичном случае
составляют более половины от общей
площади клеточных мембран (таблица 8).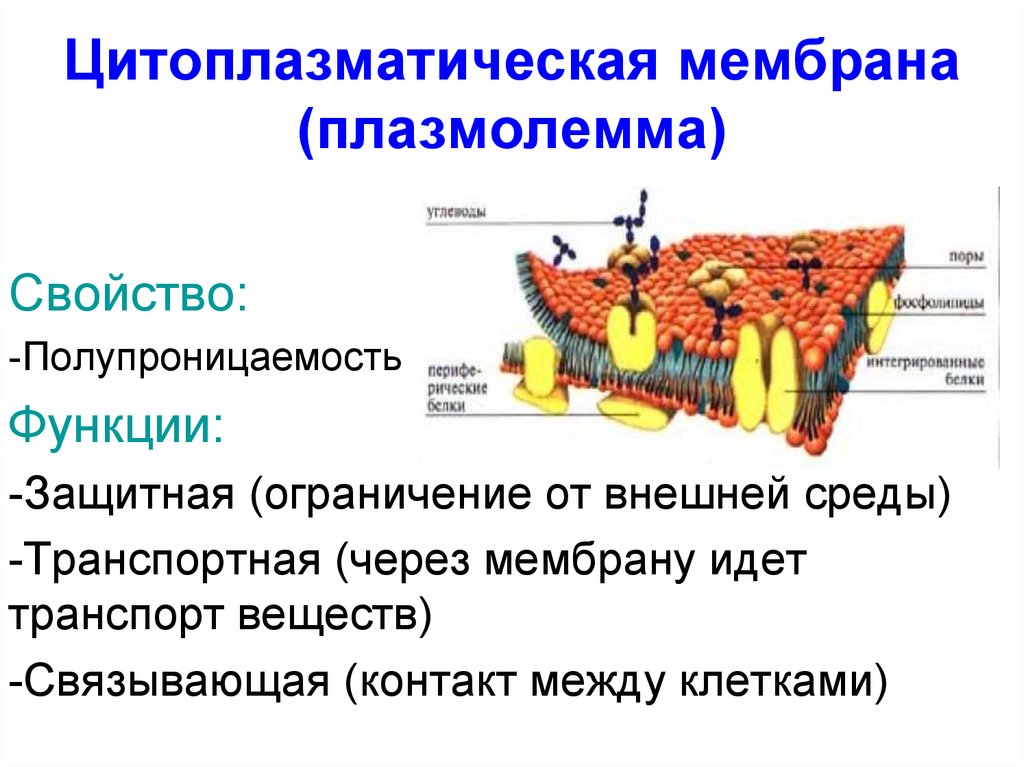
 75). Одна
диктиосома в среднем содержит 5 – 10
цистерн, хотя у низших эукариот их число
может быть больше 30. Число диктиосом на
клетку сильно варьирует в зависимости
от типа клетки – от одной до нескольких
сотен. В некоторых специализированных
клетках аппарат Гольджи может даже
занимать значительную часть объема,
хотя в типичном случае его доля невелика
(таблицы 7 и 8). Каждая отдельная цистерна
диктиосомы имеет переменную толщину:
в центре ее мембраны сближены так, что
остается просвет в 25 нм, а на периферии
имеет расширения (ампулы), ширина которых
непостоянна. От ампул отшнуровываются
многочисленные мелкие (диаметром около
50 нм) и крупные (диаметром около 1 мкм)
пузырьки (рис. 75). Эти две клетки сильно
различаются по величине: гепатоцит
имеет объем около 5000 мкм3, а секреторная
клетка поджелудочной железы – 1000 мкм3;
соответственно общин площади их клеточных
мембран оцениваются как 110 тыс. и 13 тыс.
мкм2. лизосом колеблется от 0.2 до 0.4 мкм,
а микротелец – от 0.3 до 1.5 мкм.
75). Одна
диктиосома в среднем содержит 5 – 10
цистерн, хотя у низших эукариот их число
может быть больше 30. Число диктиосом на
клетку сильно варьирует в зависимости
от типа клетки – от одной до нескольких
сотен. В некоторых специализированных
клетках аппарат Гольджи может даже
занимать значительную часть объема,
хотя в типичном случае его доля невелика
(таблицы 7 и 8). Каждая отдельная цистерна
диктиосомы имеет переменную толщину:
в центре ее мембраны сближены так, что
остается просвет в 25 нм, а на периферии
имеет расширения (ампулы), ширина которых
непостоянна. От ампул отшнуровываются
многочисленные мелкие (диаметром около
50 нм) и крупные (диаметром около 1 мкм)
пузырьки (рис. 75). Эти две клетки сильно
различаются по величине: гепатоцит
имеет объем около 5000 мкм3, а секреторная
клетка поджелудочной железы – 1000 мкм3;
соответственно общин площади их клеточных
мембран оцениваются как 110 тыс. и 13 тыс.
мкм2. лизосом колеблется от 0.2 до 0.4 мкм,
а микротелец – от 0.3 до 1.5 мкм.
 В то же время
концентрирование веществ внутри вакуоли
и полупроницаемые свойства тонопласта
способствуют тому, что вакуоль
функционирует в качестве осмометра, и
придает растительной клетке необходимую
прочность и тургисцентность (осмотическую
напряженность). Ряд веществ, накапливающихся
в вакуолях, имеет специальные
функции.
Например,
в вакуолях могут накапливаться пигменты,
придающие яркий цвет растению. Окраска
лепестков фиалки, герани или примулы
вызывается пигментами антоцианами,
накопившимися в клеточном соке вакуолей
их клеток. Другие вещества вакуолей
выполняют защитные функции. Некоторые
растения синтезируют ядовитые или
горькие вещества (алкалоиды) и накапливают
их в вакуолях. Алкалоиды высвобождаются
из ва- куолей при повреждении клеток, и
таким образом растение может защищаться
от поедания животными.
ЭПР,
аппарат Гольджи, вакуоли, лизосомы и
микротельца представляют собой единую
вакуолярную систему, поскольку их
мембраны, как будет рассказано в следующем
разделе, постоянно переходят друг в
друга в процессе функционирования этой
системы, а компоненты мембран непрерывно
совершают круговорот в клетке.
В то же время
концентрирование веществ внутри вакуоли
и полупроницаемые свойства тонопласта
способствуют тому, что вакуоль
функционирует в качестве осмометра, и
придает растительной клетке необходимую
прочность и тургисцентность (осмотическую
напряженность). Ряд веществ, накапливающихся
в вакуолях, имеет специальные
функции.
Например,
в вакуолях могут накапливаться пигменты,
придающие яркий цвет растению. Окраска
лепестков фиалки, герани или примулы
вызывается пигментами антоцианами,
накопившимися в клеточном соке вакуолей
их клеток. Другие вещества вакуолей
выполняют защитные функции. Некоторые
растения синтезируют ядовитые или
горькие вещества (алкалоиды) и накапливают
их в вакуолях. Алкалоиды высвобождаются
из ва- куолей при повреждении клеток, и
таким образом растение может защищаться
от поедания животными.
ЭПР,
аппарат Гольджи, вакуоли, лизосомы и
микротельца представляют собой единую
вакуолярную систему, поскольку их
мембраны, как будет рассказано в следующем
разделе, постоянно переходят друг в
друга в процессе функционирования этой
системы, а компоненты мембран непрерывно
совершают круговорот в клетке.
20.
Рибосома
— важнейший органоид живой клетки
сферической или слегка эллипсоидной
формы, диаметром 100—200 ангстрем, состоящий
из большой и малой субъединиц. Рибосомы
служат для биосинтеза белка из аминокислот
по заданной матрице на основе генетической
информации, предоставляемой матричной
РНК, или мРНК. Этот процесс называется
трансляцией.
В
эукариотических клетках рибосомы
располагаются на мембранах
эндоплазматического ретикулума, хотя
могут быть локализованы и в неприкрепленной
форме в цитоплазме. Нередко с одной
молекулой мРНК ассоциировано несколько
рибосом, такая структура называется
полирибосомой. Синтез рибосом у эукариот
происходит в специальной внутриядерной
структуре — ядрышке.
Схема
синтеза рибосом в клетках эукариот.
1.
Синтез мРНК рибосомных белков РНК
полимеразой II. 2. Экспорт мРНК из ядра.
3. Узнавание мРНК рибосомой и 4. синтез
рибосомных белков. 5. Синтез предшественника
рРНК (45S — предшественник) РНК полимеразой
I. 6. Синтез 5S pРНК РНК полимеразой III. 7.
 Константа
седиментации (скорость оседания в
ультрацентрифуге) рибосом эукариотических
клеток равняется 80S (большая и малая
субъединицы 60S и 40S, соответственно),
бактериальных клеток (а так же митохондрий
и пластид) — 70S (большая и малая субъединицы
50S и 30S, соответственно).
Константа
седиментации (скорость оседания в
ультрацентрифуге) рибосом эукариотических
клеток равняется 80S (большая и малая
субъединицы 60S и 40S, соответственно),
бактериальных клеток (а так же митохондрий
и пластид) — 70S (большая и малая субъединицы
50S и 30S, соответственно). Наружная
мембрана митохондрии имеет толщину
около 7 нм, не образует впячиваний и
складок, и замкнута сама на себя. На
наружную мембрану приходится около 7 %
от площади поверхности всех мембран
клеточных органелл. Основная функция —
отграничение митохондрии от цитоплазмы.
Наружная мембрана митохондрии состоит
из билипидного слоя и пронизывающих
его белков; соотношение липидов и белков
по массе — примерно 1:1. Особую роль
играет порин — каналообразующий белок:
он формирует в наружной мембране
отверстия диаметром 2-3 нм, через которые
могут проникать небольшие молекулы и
ионы весом до 5 кДа.
Крупные молекулы могут пересекать
наружную мембрану только посредством
активного транспорта через транспортные
белки митохондриальных мембран.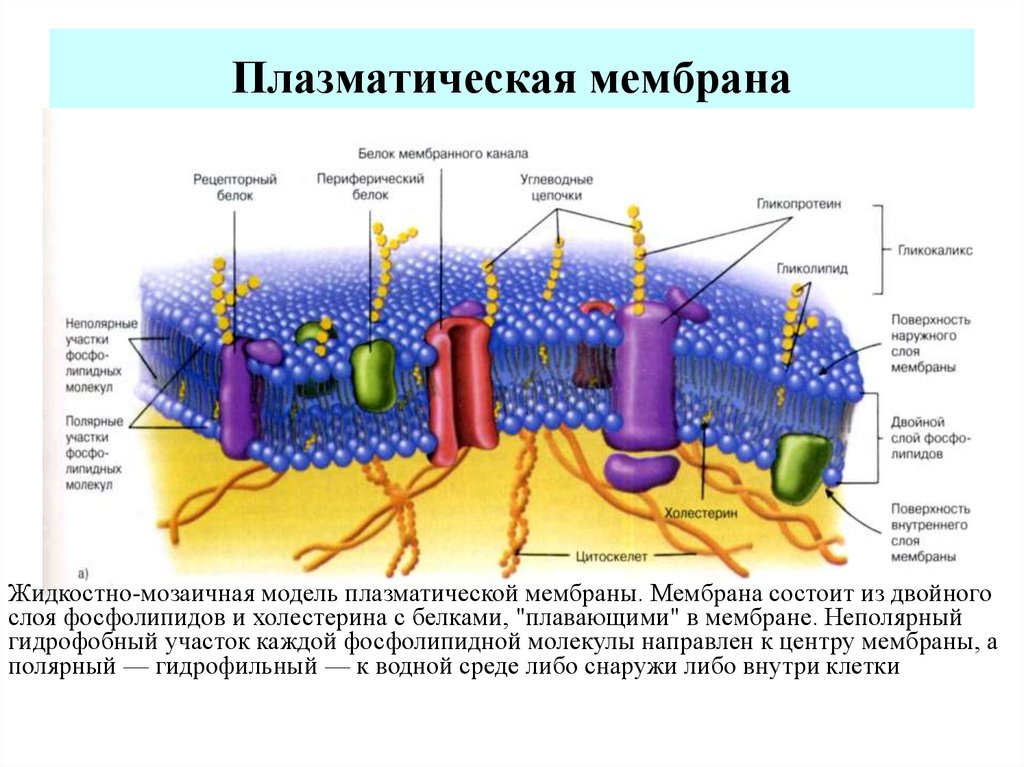 Для
наружной мембраны характерно
присутствие ферментов:
монооксигеназы, ацил-СоА-синтетазы и
фосфолипазы А2.
Наружная мембрана митохондрии может
взаимодействовать с мембраной эндоплазматического
ретикулума;
это играет важную роль в транспортировке
липидов и ионов кальция.
Для
наружной мембраны характерно
присутствие ферментов:
монооксигеназы, ацил-СоА-синтетазы и
фосфолипазы А2.
Наружная мембрана митохондрии может
взаимодействовать с мембраной эндоплазматического
ретикулума;
это играет важную роль в транспортировке
липидов и ионов кальция.
Межмембранное пространство
Межмембранное пространство представляет собой пространство между наружной и внутренней мембранами митохондрии. Его толщина — 10-20 нм. Так как наружная мембрана митохондрии проницаема для небольших молекул и ионов, их концентрацияв периплазматическом пространстве мало отличается от таковой в цитоплазме. Напротив, крупным белкам для транспорта из цитоплазмы в периплазматическое пространство необходимо иметь специфические сигнальные пептиды; поэтому белковые компоненты периплазматического пространства и цитоплазмы различны. Одним из белков, содержащихся в периплазматическом пространстве, является цитохром c — один из компонентов дыхательной цепи митохондрий.
Внутренняя мембрана
Внутренняя
мембрана образует многочисленные
гребневидные складки — кристы,
существенно увеличивающие площадь ее
поверхности и, например, в
клетках печени составляет
около трети всех клеточных мембран. Характерной чертой состава внутренней
мембраны митохондрий является присутствие
в ней кардиолипина —
особого фосфолипида,
содержащего сразу четыре жирные
кислоты и
делающего мембрану абсолютно непроницаемой
для протонов.
Ещё одна особенность внутренней мембраны
митохондрий — очень высокое
содержание белков (до
70 % по весу), представленных транспортными
белками,ферментами дыхательной
цепи,
а также крупными АТФ-синтетазными комплексами.
Внутренняя мембрана митохондрии в
отличие от внешней не имеет специальных
отверстий для транспорта мелких молекул
и ионов; на ней, на стороне, обращенной
к матриксу, располагаются особые
молекулы АТФ-синтазы,
состоящие из головки, ножки и основания.
При прохождении через них протонов происходит
синтез АТФ.
В основании частиц, заполняя собой всю
толщу мембраны, располагаются
компонентыдыхательной
цепи.
Наружная и внутренняя мембраны в
некоторых местах соприкасаются, там
находится специальный белок-рецептор,
способствующий транспорту митохондриальных
белков, закодированных в ядре, в матрикс
митохондрии.
Характерной чертой состава внутренней
мембраны митохондрий является присутствие
в ней кардиолипина —
особого фосфолипида,
содержащего сразу четыре жирные
кислоты и
делающего мембрану абсолютно непроницаемой
для протонов.
Ещё одна особенность внутренней мембраны
митохондрий — очень высокое
содержание белков (до
70 % по весу), представленных транспортными
белками,ферментами дыхательной
цепи,
а также крупными АТФ-синтетазными комплексами.
Внутренняя мембрана митохондрии в
отличие от внешней не имеет специальных
отверстий для транспорта мелких молекул
и ионов; на ней, на стороне, обращенной
к матриксу, располагаются особые
молекулы АТФ-синтазы,
состоящие из головки, ножки и основания.
При прохождении через них протонов происходит
синтез АТФ.
В основании частиц, заполняя собой всю
толщу мембраны, располагаются
компонентыдыхательной
цепи.
Наружная и внутренняя мембраны в
некоторых местах соприкасаются, там
находится специальный белок-рецептор,
способствующий транспорту митохондриальных
белков, закодированных в ядре, в матрикс
митохондрии.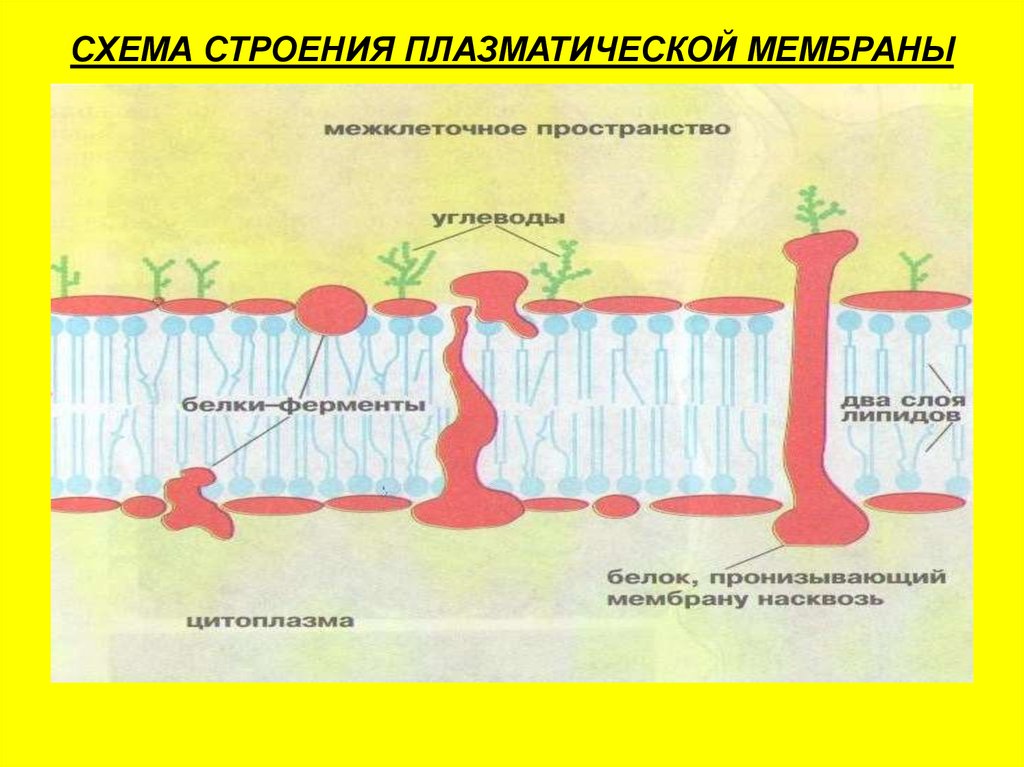
Матрикс
Матрикс — ограниченное внутренней мембраной пространство. В матриксе (розовом веществе) митохондрии находятся ферментные системы окисления пирувата жирных кислот, а также ферменты цикла трикарбоновых кислот (цикла Кребса). Кроме того, здесь же находится митохондриальная ДНК, РНК и собственный белоксинтезирующий аппарат митохондрии.
4.4B: Наружная мембрана грамотрицательных бактерий — Биология LibreTexts
- Последнее обновление
- Сохранить как PDF
- Идентификатор страницы
- 8844
- Безграничный
- Безграничный
Цели обучения
- Распознать характеристики грамотрицательных бактерий
Грамотрицательная клеточная стенка состоит из наружной мембраны, слоя пептидоглигана и периплазмы .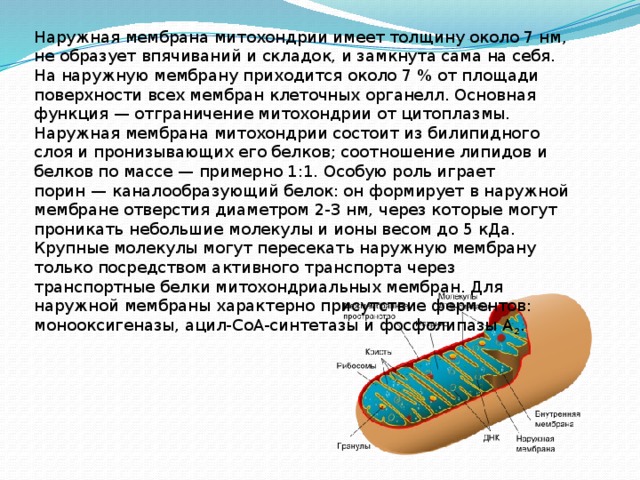
У грамотрицательных бактерий клеточная стенка состоит из одного слоя пептидогликана, окруженного мембранной структурой, называемой внешней мембраной. Грамотрицательные бактерии не сохраняют кристаллический фиолетовый, но способны сохранять контрастный краситель, обычно сафранин, который добавляют после кристаллического фиолетового. Сафранин отвечает за красный или розовый цвет, наблюдаемый у грамотрицательных бактерий. Клеточная стенка грамотрицательных бактерий тоньше (толщиной 10 нанометров) и менее плотная, чем у грамположительных бактерий, но остается прочной, жесткой и эластичной, что придает им форму и защищает от экстремальных условий окружающей среды. Наружная мембрана грамотрицательных бактерий неизменно содержит уникальный компонент — липополисахарид (ЛПС) в дополнение к белкам и фосфолипидам. Молекула LPS токсична и классифицируется как эндотоксин, который вызывает сильный иммунный ответ, когда бактерии заражают животных.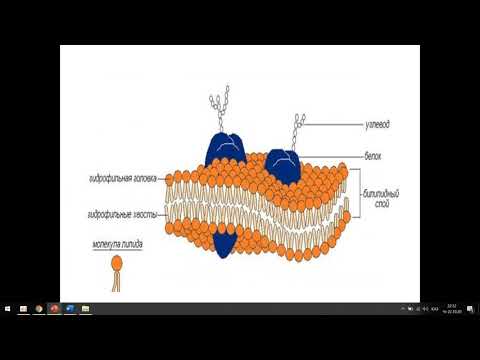
У грамотрицательных бактерий наружная мембрана обычно рассматривается как часть внешнего листка мембранной структуры и относительно проницаема. Он содержит структуры, которые помогают бактериям прикрепляться к клеткам животных и вызывать заболевания. Слой пептидогликана нековалентно связан с молекулами липопротеинов, называемыми липопротеинами Брауна, через их гидрофобную головку. Между внешней мембраной и плазматической мембраной в периплазматическом пространстве находится концентрированный гелеобразный матрикс (периплазма). Фактически это неотъемлемая часть стенки грамотрицательной клетки, содержащая белки, связывающие аминокислоты, сахара, витамины, железо и ферменты, необходимые для питания бактерий. Периплазматическое пространство может действовать как резервуар для факторов вирулентности и динамического потока макромолекул, отражающих метаболический статус клетки и ее реакцию на факторы окружающей среды. Вместе плазматическая мембрана и клеточная стенка (внешняя мембрана, пептидогликановый слой и периплазма) составляют грамотрицательную оболочку.
Ключевые моменты
- Наружная мембрана грамотрицательных бактерий содержит липополисахариды, белки и фосфолипиды.
- Липополисахаридный компонент действует как фактор вирулентности и вызывает заболевание у животных.
- В периплазматическом пространстве между наружной мембраной и плазматической мембраной содержится больше факторов вирулентности.
Ключевые термины
- липополисахарид : любой из большого класса липидов, конъюгированных с полисахаридами
- эндотоксин : Любой токсин, выделяемый микроорганизмом и высвобождаемый в окружающую среду только после его гибели.
Эта страница под названием 4.4B: Gram-Negative Outer Membrane распространяется по лицензии CC BY-SA 4.0 и была создана, изменена и/или курирована Boundless.
- Наверх
- Была ли эта статья полезной?
- Тип изделия
- Раздел или Страница
- Автор
- Безграничный
- Лицензия
- CC BY-SA
- Версия лицензии
- 4,0
- Показать оглавление
- нет
- Теги
- Мембрана (грамотрицательная)
- пептидоглиган
- периплазма
Белки наружной мембраны | SimBac – моделирование бактериальных систем
Схематические различия между грамположительными (слева) и грамотрицательными (справа) бактериями.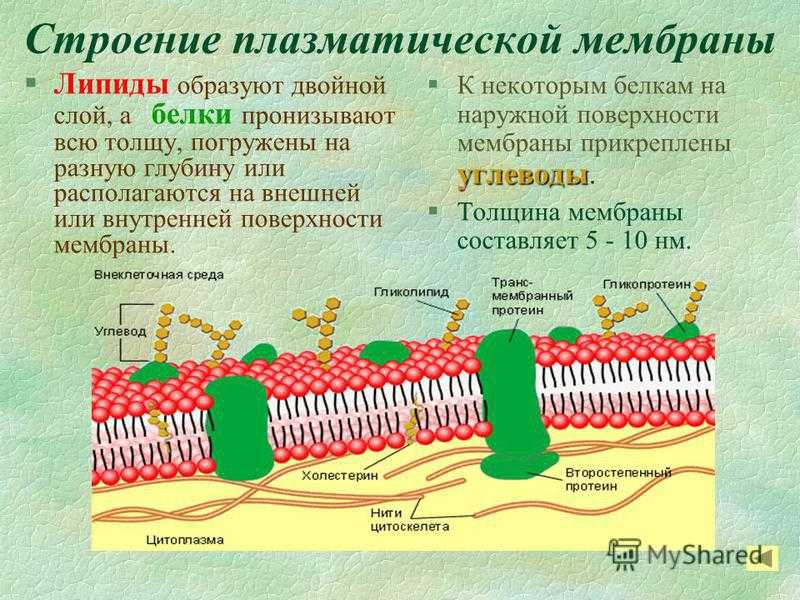 Первые имеют один бислой с окутывающей пептидогликановой клеточной стенкой, защищающей бактерию, тогда как вторые имеют две клеточные мембраны, разделенные областью периплазмы, внутри которой находится клеточная стенка.
Первые имеют один бислой с окутывающей пептидогликановой клеточной стенкой, защищающей бактерию, тогда как вторые имеют две клеточные мембраны, разделенные областью периплазмы, внутри которой находится клеточная стенка.
Наружная мембрана грамотрицательных бактерий (обладающих двумя оболочечными мембранами с периплазмой между ними, как показано вверху справа) уникальна по ряду причин. Прежде всего, это состав внешней мембраны. В то время как его внутренний листок содержит типичные двусторонние фосфолипиды, внешний листок состоит из липополисахаридов (ЛПС). Молекулы ЛПС имеют 5-6 ацильных хвостов, соединенных с цепочкой молекул сахара, простирающейся над мембраной (см. рисунок ниже). Некоторые виды грамотрицательных бактерий также могут содержать цепи О-антигена, выступающие из ядра олигосахарида.
(A) пример молекулы ЛПС. Состав внутреннего и внешнего ядра варьируется внутри и между видами (B) стандартный амфифильный двойной слой фосфолипидов. (C) модель внешней мембраны с LPS во внешней (вверху) и фосфолипидами во внутренней (внизу) листочках.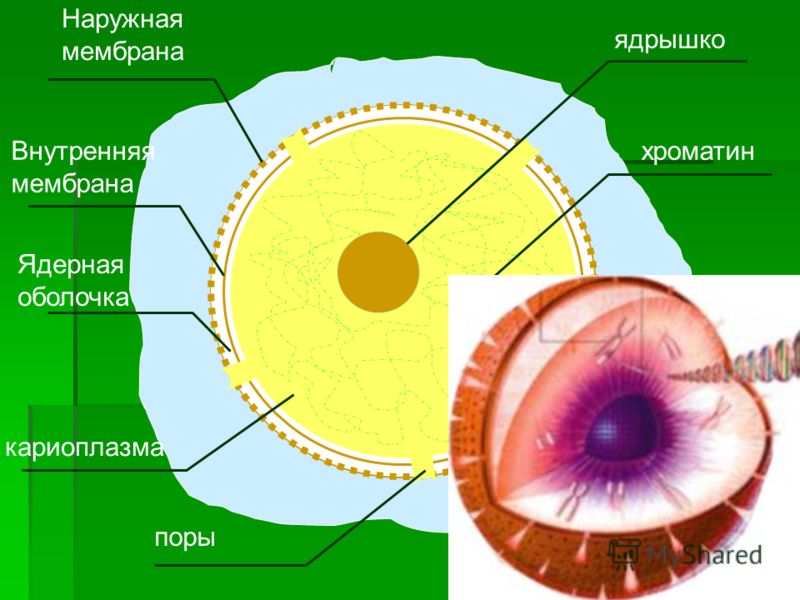
TonB (красный) «тянет» люменальный домен транспортера BtuB (синий), соединение опосредовано водородными связями (желтый).
Другим уникальным аспектом внешней мембраны является то, что она полна поринов, которые обеспечивают относительно легкую диффузию небольших молекул через нее. Таким образом, энергия не может быть произведена или сохранена здесь. Тем не менее, бактериям часто необходимо активно импортировать большие и/или дефицитные питательные вещества через внешнюю мембрану. Чтобы получить энергию для этой задачи, они используют белок внутренней мембраны, TonB, который соединяется через периплазму с переносчиком внешней мембраны для импорта, например, витамина B12 или железа, извлеченного из трансферрина человека. Механизмы этого взаимодействия и то, как оно вызывает транспорт, все еще остаются открытыми вопросами в области микробиологии. Мы предположили, что энергия передается через механическое соединение между комплексами внутренней и внешней мембран, что подтверждается моделированием, демонстрирующим прочность соединения TonB-транспортер, опосредованного водородными связями, а также реакцию транспортера на приложение силы.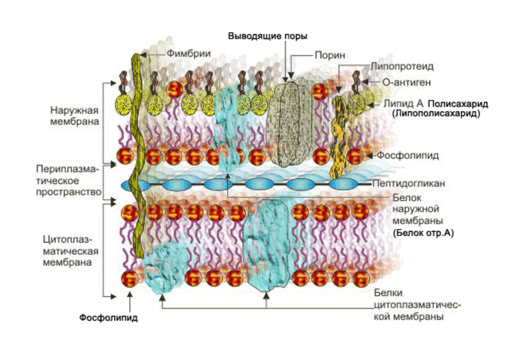 . В частности, мы видим во многих контекстах, что внутренний домен транспортера разворачивается, создавая путь для субстрата через мембрану.
. В частности, мы видим во многих контекстах, что внутренний домен транспортера разворачивается, создавая путь для субстрата через мембрану.
Модель полноразмерного автовоза ЭспП.
Так же, как бактериям необходимо импортировать молекулы через внешнюю мембрану, иногда им необходимо их экспортировать. Например, некоторые факторы вирулентности секретируются с помощью так называемых «автотранспортеров», белков, которые экспортируют сами себя! Автотранспортер состоит из двух доменов: связанного с мембраной ствола на С-конце и секретируемого домена на N-конце. После транспорта N-концевой домен обычно саморасщепляется, освобождая его от мембраны. Мы провели моделирование нативного предварительно расщепленного состояния, а также на ряде биохимически охарактеризованных мутантов. Моделирование выявило необычную конформацию каталитического остатка аспарагина, наблюдаемую менее чем в 2% всех атомных структур, которая поддерживается уникальной электростатической средой, создаваемой соседними остатками.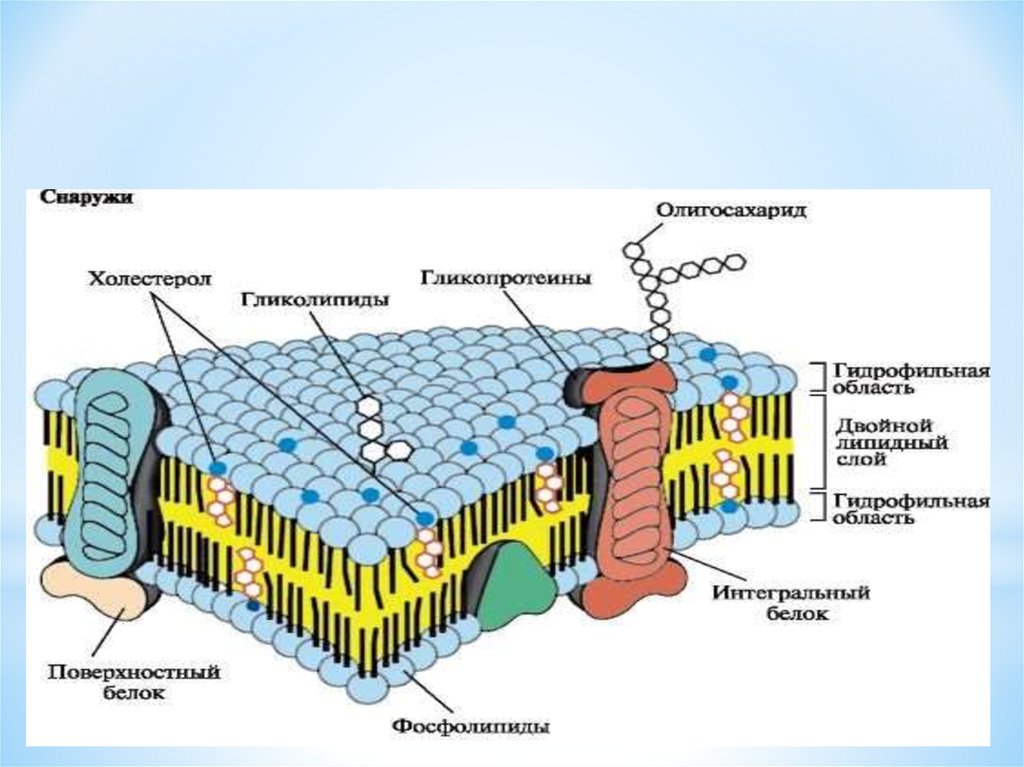 Предполагается, что эта конформация значительно ускорит реакцию расщепления, а другие обнаруженные вовлеченные остатки согласуются с экспериментальными наблюдениями.
Предполагается, что эта конформация значительно ускорит реакцию расщепления, а другие обнаруженные вовлеченные остатки согласуются с экспериментальными наблюдениями.
Публикации
- Структурные основы пиратства железа патогенными Neisseria .
Н. Ноинадж, Н. К. Исли, М. Оке, Н. Мизуно, Дж. Гумбарт , Э. Бура, А. Н. Стир, О. Зак, П. Айсен, Э. Тайхоршид, Р. В. Эванс, А. Р. Горриндж, А. Б. Мейсон, А. К. Стивен и С. К. Бьюкенен. Природа , 483:53-58, 2012. - Молекулярная основа для активации каталитического остатка аспарагина в саморасщепляющемся бактериальном аутотранспортере.
Т. Дж. Барнард, Джеймс Гамбарт , Дж. Х. Петерсон, Н. Ноинадж, Н. К. Исли, Н. Даутин, А. Дж. Кушак, Э. Тайхоршид, Х. Д. Бернстайн и С. К. Бьюкенен. Журнал молекулярной биологии , 415:128-142, 2012. - Соединение связывания кальция и субстрата посредством выравнивания петель в переносчике наружной мембраны BtuB.
