Мембраны: немного физики | BASK
Статья о физических основах работы мембранных тканей. Процессы, происходящие в поровых и непоровых мембранах, имеют разную природу. Отсюда их отличия, сильные и слабые стороны.
Высокие технологии, изначально работавшие в космосе или на сложном производстве, со временем становятся повседневными. Так, мембранные материалы, несколько десятилетий назад родившиеся в научных лабораториях, теперь служат нам в одежде и обуви.
Мембрана – это барьер, который пропускает через себя одни вещества и останавливает другие. Впервые применить это свойство мембран в одежде догадались в фирме Gore в 70-хх годах прошлого века. Именно в это время стало технологически возможным создать прочную поровую мембрану на основе полимеров. Радиус пор подбирался так, чтобы они не пропускали капли воды из внешней среды и не создавали препятствия для выхода пара наружу.
Капля тумана – 0,1 мм (100000 нм)
Крупная капля дождя – 6,5 мм (6500000 нм)
Молекула воды – 0,2 нм
d пор в мембране = 140 – 200 нм
Зачем это нужно?
Простой полиэтиленовый плащ отлично защитит от дождя и ветра, но не спасёт от намокания. А всё потому, что одной из важнейших постоянных человеческого тела является температура. Она может колебаться в узких пределах. При любой активности мы вырабатываем тепло, излишки которого незамедлительно сбрасываются в окружающую среду. Одним из механизмов отвода тепла является потоотделение: ежесуточно человек выделяет не менее полулитра пота в нормальных условиях, при высоких нагрузках и температурах это значение достигает 12 литров.
А всё потому, что одной из важнейших постоянных человеческого тела является температура. Она может колебаться в узких пределах. При любой активности мы вырабатываем тепло, излишки которого незамедлительно сбрасываются в окружающую среду. Одним из механизмов отвода тепла является потоотделение: ежесуточно человек выделяет не менее полулитра пота в нормальных условиях, при высоких нагрузках и температурах это значение достигает 12 литров.
Но полиэтилен препятствует отводу паров воды, и она конденсируется на его поверхности, опять превращаясь в воду – порочный круг замыкается, под плащом пасмурно и сыро, особенно если снаружи тепло и солнечно.
Как работают поровые мембраны?
Поровая мембрана позволяет избежать «застоя» паров воды под одеждой.
Водяной пар – газообразное состояние той же воды, что падает на нас снаружи в жидком состоянии в виде капель.
Расстояние между молекулами воды в паре так велико, что каждая молекула выступает как отдельный «игрок»: поровая мембрана для неё не является серьёзным препятствием и молекула свободно выходит наружу. Движение пара в поровой мембране можно описать как поток через отверстия.
Движение пара в поровой мембране можно описать как поток через отверстия.
В капле те же молекулы воды связаны силой поверхностного натяжения, они живут плотными сообществами. Такой конгломерат не может разом пройти сквозь пору пока не растечётся по поверхности монослоем в размер молекулы. В этот момент мембрана начинает пропускать воду. Чтобы предотвратить такой ход событий на поверхность мембраны наносят специальное гидрофобное покрытие с плохой смачиваемостью – DWR. С этим покрытием молекулы воды не могут прореагировать и капля не растекаясь скатывается с поверхности. Да и сама пористая мембрана производится из гидрофобных полимеров, например, политетрафторэтилена, чтобы у молекул пара не было возможности остаться на стенке поры по пути наружу.
Политетрафторэтилен (ПТФЭ) – превосходит по химической стойкости все полимеры: не смачивается водой и не растворяется в обычных растворителях. Полимер с самой большой молекулярной массой.
Почему молекулы пара начинают целенаправленно двигаться наружу? Что их подталкивает?
Всё мире стремится к равновесию, по крайне мере, так говорит физика.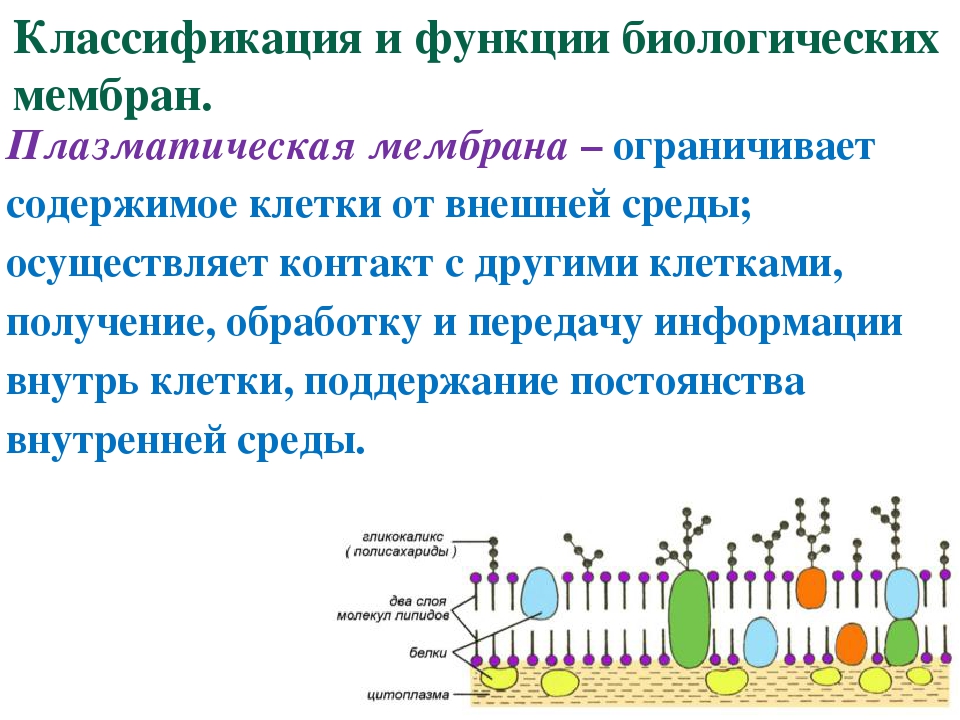 Любое движение обусловлено избытком энергии/вещества в одном месте и недостатком в другом. Когда парциальное давление водяного пара под мембраной (под курткой) превышает его парциальное давление во внешней среде, молекулы воды начинают двигаться сквозь мембрану наружу. Процесс останавливается, когда давление пара внутри и снаружи выравнивается.
Любое движение обусловлено избытком энергии/вещества в одном месте и недостатком в другом. Когда парциальное давление водяного пара под мембраной (под курткой) превышает его парциальное давление во внешней среде, молекулы воды начинают двигаться сквозь мембрану наружу. Процесс останавливается, когда давление пара внутри и снаружи выравнивается.
Значит, если снаружи влажно, то отвод водяного пара прекращается, и мы промокаем? Да, но только в случае, когда температуры под курткой и снаружи близки по значениям. Если стоит холодная влажная погода, то отвод водяного пара будет продолжаться даже в таких условиях, так как парциальное давление зависит от температуры, и разница в давлениях сохранится.
Особенности поровых мембран
Рабочие характеристики поровой мембраны в первую очередь определяются количеством пор, их размером, распределением по поверхности и толщиной мембраны, а не химическими свойствами материала, из которого она изготовлена. Со временем на поверхности мембраны появляются грязевые отложения, они сильно снижают её качество, закупоривая поры и меняя свойства. Неправильно подобранное моющее средство, например порошок, также забивает поры. Поэтому уход за поровыми мембранами требует особого внимания и осторожности. А гидрофобность внешней ткани – основа правильной работы этого сложного механизма, поддерживается при помощи пропиток.
Неправильно подобранное моющее средство, например порошок, также забивает поры. Поэтому уход за поровыми мембранами требует особого внимания и осторожности. А гидрофобность внешней ткани – основа правильной работы этого сложного механизма, поддерживается при помощи пропиток.
Непоровые мембраны. Зачем нужны они?
Непоровые мембраны – отдельный класс мембран, работа которых основана на совершенно иных физических процессах. Как следует из названия, в них нет пор, однако материал мембраны имеет неравномерную структуру, причина которой – свободный объём между большими молекулами полимера. Этот объём достаточен для движения небольших молекул воды по мембране – диффузии. Молекулярный размер этих полостей не сравним с размерами пор.
Водяной пар химически связывается с гидрофильной внутренней поверхностью мембраны. В случае интенсивной работы с повышенным потоотделением влага конденсируется на ней. Материал мембраны, обычно полиуретан, начинает набухать, впитывая в себя молекулы воды. Все молекулы под действием температуры начинают двигаться быстрее: гибкий, за счёт влаги, полимер расширяется, пространство между цепочками увеличивается и диффузия молекул воды ускоряется. Она происходит до тех пор, пока концентрация молекул воды в мембране не станет одинаковой по всей толщине мембраны. Основными движущими силами процессов в непоровой мембране являются: разница концентрации молекул воды в ней и разница температур с двух сторон от мембраны. Движение направлено из области с высокой концентрацией воды в область с низкой концентрацией воды. Молекулы, проходящие через свободные полости в мембране, достигают наружней поверхности и уходят в окружающую среду.
Все молекулы под действием температуры начинают двигаться быстрее: гибкий, за счёт влаги, полимер расширяется, пространство между цепочками увеличивается и диффузия молекул воды ускоряется. Она происходит до тех пор, пока концентрация молекул воды в мембране не станет одинаковой по всей толщине мембраны. Основными движущими силами процессов в непоровой мембране являются: разница концентрации молекул воды в ней и разница температур с двух сторон от мембраны. Движение направлено из области с высокой концентрацией воды в область с низкой концентрацией воды. Молекулы, проходящие через свободные полости в мембране, достигают наружней поверхности и уходят в окружающую среду.
Внешняя поверхность непоровой мембраны всегда защищена гидрофобной тканью с плотным плетением, которая не пропускает капли воды снаружи и не препятствует отводу пара.
Полиуретан – прочный, устойчивый к старению, эластичный материал. Гидрофилен. Плёнки из полиуретана легко наносятся на поверхности любой конфигурации.Обладает относительно низкой молекулярной массой.
Слабые и сильные стороны
Непоровая мембрана меньше загрязняется, так как в ней нет пор и нечему забиваться. Эластичность этого полимера даёт возможность производить тянущиеся ткани на его основе.
Проблемным местом непоровой мембраны нужно назвать ограничение её работы при низких температурах. Влага на внутренней поверхности мембраны превращается в изморозь, это затрудняет отведение пара и причиняет дискомфорт. В случае с поровой мембраной изморозь не во всех случаях образуется на внутренней стороне мембраны, так как точка росы может оказаться в другом месте за счёт интенсивного потока пара через мембрану.
Тесты показывают, что поровая мембрана более устойчива к образованию конденсата на внутренней поверхности и лучше работает в условиях повышенной влажности – JIS L 1099 A1.
Непоровая мембрана начинает работать с небольшой задержкой: ей требуется время на набухание (намокание), чтобы выйти на максимальные рабочие показатели. Тесты показывают, что эффективность непоровой мембраны возрастает с ростом абсолютной влажности под одеждой, поровая мембрана мало зависит от этого параметра. В условиях активной работы при невысокой влажности окружающего воздуха, хорошие непоровые мембраны обходят поровые по паропроницаемости – JIS L 1099 B1.
Тесты показывают, что эффективность непоровой мембраны возрастает с ростом абсолютной влажности под одеждой, поровая мембрана мало зависит от этого параметра. В условиях активной работы при невысокой влажности окружающего воздуха, хорошие непоровые мембраны обходят поровые по паропроницаемости – JIS L 1099 B1.
Из-за гидрофильности самой мембраны её водостойкость ниже, чем у полностью гидрофобной пористой. Когда верхний защитный слой ткани с непоровой мембраной начинает пропускать влагу, она мгновенно проникает внутрь.
Комбинированные мембраны
Хорошие результаты часто получают на стыке технологий. В последнее время всё больше интереса вызывают комбинированные мембраны, которые включают поровую мембрану у наружнего слоя и тонкую непоровую со стороны тела. Так производители защищают пористую мембрану от загрязнений тела и увеличивают показатели паропроницаемости.
Так же интересен опыт создания поровой мембраны из волокон полиуретана, реализованный в тканях Polartec® NeoShell®.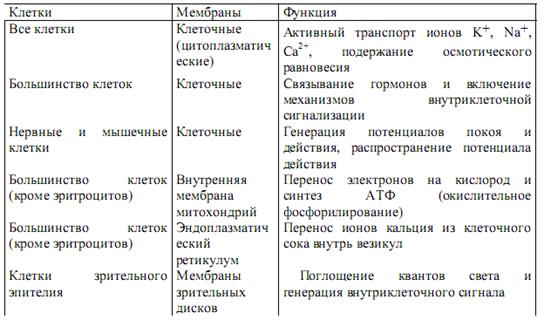
Характеристики мембран
- Влагостойкость
- Паропроницаемость (A1, B1)
- Ret
Поровые мембраны в снаряжении BASK:
Polartec® Neoshell
Advance® Alaska
Advance® Perfomance
Advance® 2.5L
Непоровые мембраны в снаряжении BASK:
Gelanots®
Dermizax® 3L
Ключ от клетки: как открыть живые ворота мембраны
Липидная мембрана клетки — основа клеточной оболочки любого живого организма — это удивительный умный «забор», через который клетка общается с организмом, питается, дышит, защищается от вторжения интервентов и чужаков, впускает нужные вещества и закрывается от нежелательных. Это целый комплекс security-мер с избирательным воздействием. Основной инструмент этой биохимической «коммуникации» — поры, опциональные отверстия в мембране. Своеобразный пропускной шлюз, который ученые активно изучают и описывают, чтобы в дальнейшем управлять им в собственных — благих, разумеется, целях.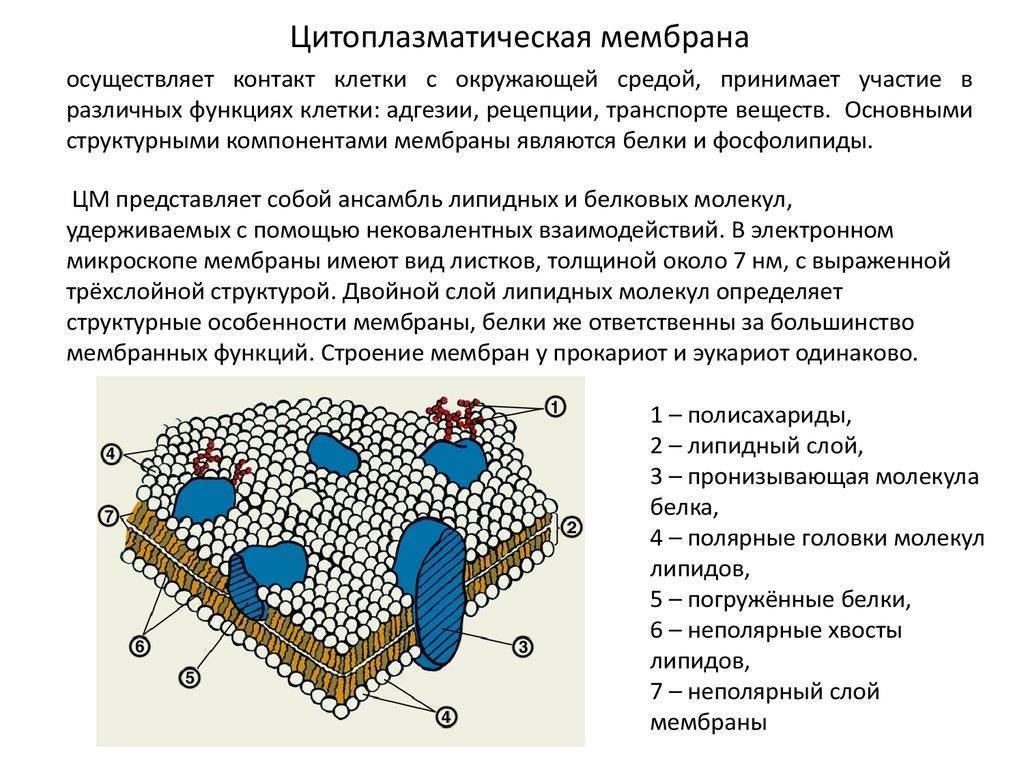
В чем суть исследования и что сделано
Ученые впервые полностью описали процесс образования пор в липидных мембранах и осуществили компьютерное моделирование их образования и эволюции. Они создали масштабную теоретическую модель, которая объяснила несостыковки в полученных ранее экспериментальных данных других исследований и разрешила накопившиеся противоречия.
Результаты работы коллаборации ученых из НИТУ «МИСиС», Института физической химии и электрохимии имени А.Н. Фрумкина РАН и Института биоорганической химии имени академиков М.М. Шемякина и Ю.А. Овчинникова РАН под руководством Олега Батищева опубликована в двух частях в журнале Scientific Reports: первая, вторая.
Липидные мембраны — это оболочки, которые отделяют клетки и их органеллы от внешней среды. Эти структуры выполняют ряд важных функций жизнедеятельности, в частности становятся барьером, который контролирует обмен веществ клетки. Возможные нарушения этого барьерного механизма давно и активно изучаются в свете разработки лекарств и терапевтических стратегий, таких, как доставка препаратов, поскольку именно мембрана в конечном счете решает и определяет, попадет ли то или иное вещество в клетку. Соответственно, алгоритм «правильного» попадания вещества через мембрану путем создания поры — это и есть ID-карта в живую клетку.
Соответственно, алгоритм «правильного» попадания вещества через мембрану путем создания поры — это и есть ID-карта в живую клетку.
Несмотря на то, что в мире существует множество экспериментально проверенных методов создания в мембране пор, через которые препарат может проникнуть в клетку (например, антибиотик, чтобы убивать бактерии или антиопухолевый токсин, чтобы уничтожать клетки рака), до сих пор не было физической модели, которая описывает формирование, рост и устойчивость таких пор.
Как сделано
Авторы задались целью создать полную теоретическую модель, которая бы описывала все стадии эволюции поры в липидной мембране. Эта задача осложняется тем, что любые попытки представить мембрану в качестве идеальной упругой оболочки без учета особенностей внутреннего строения живого «забора» приводили лишь к упрощенному и потому очень грубому описанию этой системы. Чтобы устранить подобные проблемы, ученые начали с максимально полного теоретического описания мембраны, а затем при помощи ряда преобразований получили выражения для энергии поры, позволяющее описать состояние поры в зависимости от ее геометрических параметров.
С помощью новой компьютерной модели ученые смогли объяснить несостыковки, наблюдавшиеся во многих работах, посвящённых данной тематике. Эта модель не только объясняет сам механизм возникновения пор в мембране, с ее помощью можно заранее описать, как именно мембрана отреагирует на механическое (укол, прокол) или электромагнитное воздействие (точечное облучение полем): в некоторых случаях оно приводит к управляемому формированию поры определенных размеров, а в некоторых — к необратимому разрыву мембраны и гибели клетки. Этот вариант, разумеется, нужно исключить в случае терапии, и наоборот — можно широко использовать для непосредственного устранения зараженных клеток.
Для того чтобы окончательно убедиться в справедливости выдвинутой теории, ученые также провели компьютерное моделирование методами молекулярной динамики, в котором липидная мембрана воссоздавалась на масштабе отдельных молекул. Результаты этих исследований хорошо совпадали с предсказанием теоретической модели и имеющимися экспериментальными данными, а также позволили наглядно «увидеть», как эволюционирует (возникает, растет и расширяется) пора в виртуальной мембране.
Рассказывает соавтор статьи, научный сотрудник кафедры теоретической физики и квантовых технологий НИТУ «МИСиС» Тимур Галимзянов:
«Эта работа потребовала очень больших трудозатрат от всех участников проекта, большого объема машинного времени для расчётов методами молекулярной динамики, проведённых коллегами из лаборатории моделирования биомолекулярных систем ИБХ РАН; долгой работы по построению моделей наблюдаемых процессов; и, главное проведения огромного массива расчетов, во многом аналитических, выполненных, в основном, Сергеем Акимовым, сотрудником ИФХЭ РАН и кафедры теоретической физики и квантовых технологий НИТУ «МИСиС».
Зачем сделано
Авторы надеются, что их работа станет фундаментом для будущих исследований, посвященных контролируемой доставке различных препаратов в клетку. Грубо говоря, компьютерная модель сложной органической системы — липидной мембраны — поможет подбирать оптимальные режимы воздействия на нее для успешного прохода через «шлюз» клетки в обход всех security-мер и введения внутрь нужных концентраций нужных веществ. Кроме того, новая модель, вероятно, поможет описать процессы, связанные с нарушением целостности мембран, что наблюдается в ходе многих сложных и пока не поддающихся лечению нейродегенеративных заболеваний, таких как болезни Альцгеймера, Паркинсона, Пика, хореи Гентингтона и тд.
Кроме того, новая модель, вероятно, поможет описать процессы, связанные с нарушением целостности мембран, что наблюдается в ходе многих сложных и пока не поддающихся лечению нейродегенеративных заболеваний, таких как болезни Альцгеймера, Паркинсона, Пика, хореи Гентингтона и тд.
«Никогда раньше мы не проводили таких подробных и последовательных теоретических исследований. Их результат полностью оправдал потраченные усилия: нам впервые удалось построить полную модель процесса формирования пор в мембранах, позволяющую делать не только качественные, но и количественные предсказания».
Мягкие мембраны и битумная черепица кровель – особенности применения кабельных антиобледенительных систем
Введение
Среди множества типов кровельных покрытий существуют такие, которые привлекают пользователей своей практичностью и привлекательным внешним видом. Это — битумная черепица, катепал, ондулин и т. п. Кроме того, при проектировании плоских кровель в настоящее время незаменимым покрытием стали мягкие мембраны. Такие покрытия при правильной эксплуатации гарантированно долговечны и полностью исключают протечки. В отличие от металлических кровель (листовые железо или медь, металлочерепица и пр.) эти покрытия прекрасно гасят шум дождя, чем привлекают многих частных пользователей. Однако, в странах со снежными зимами эксплуатация таких крыш сопряжена с необходимостью борьбы с наледью и снегом. А причиной этого является промерзание водоприёмных воронок внутренних водосливов на плоских мембранных кровлях или плохое скольжение снега на покрытиях с каменной крошкой (результат — значительное скопление снега). Механическая очистка таких кровель от снега и льда практически исключена. Велик риск повреждения кровли. Однако беспроблемная эксплуатация таких кровельных покрытий возможна благодаря разработанным и хорошо себя зарекомендовавшим на практике антиобледенительным системам (АОС) DEVI.
Такие покрытия при правильной эксплуатации гарантированно долговечны и полностью исключают протечки. В отличие от металлических кровель (листовые железо или медь, металлочерепица и пр.) эти покрытия прекрасно гасят шум дождя, чем привлекают многих частных пользователей. Однако, в странах со снежными зимами эксплуатация таких крыш сопряжена с необходимостью борьбы с наледью и снегом. А причиной этого является промерзание водоприёмных воронок внутренних водосливов на плоских мембранных кровлях или плохое скольжение снега на покрытиях с каменной крошкой (результат — значительное скопление снега). Механическая очистка таких кровель от снега и льда практически исключена. Велик риск повреждения кровли. Однако беспроблемная эксплуатация таких кровельных покрытий возможна благодаря разработанным и хорошо себя зарекомендовавшим на практике антиобледенительным системам (АОС) DEVI.
В настоящей статье мы рассмотрим особенности проектирования, выбора оборудования и способы крепления нагревательных кабелей на мембранах и битумносодержащих покрытиях кровель.
Проектирование антиобледенительной системы
Настенные желоба, карнизы, ендовы
Для хорошо теплоизолированных крыш, то есть выполненных в соответствии с современными строительными требованиями, необходимая линейная мощность обогрева зон, прилегающих к настенным желобам, на карнизах или в водосборных ендовах, отводящих талую воду в водосливы, составляет 40…50 Вт/м. Если крыша имеет плохую теплоизоляцию, что характерно для мансардных этажей зданий, этот параметр должен быть больше: 50…60 Вт/м. Имеется в виду мощность нагревательного кабеля, расположенного в зоне обогрева желобов, карнизов и ендов на длине 1 метр. Рекомендуемый шаг укладки саморегулируемого или резистивного нагревательного кабеля с постоянной линейной мощностью около 20 Вт/м составляет 7,5 см для хорошо теплоизолированных крыш и 5 см для крыш с большим поступлением тепла от жилых помещений, технических устройств, расположенных на крыше и от солнца на южных скатах кровли.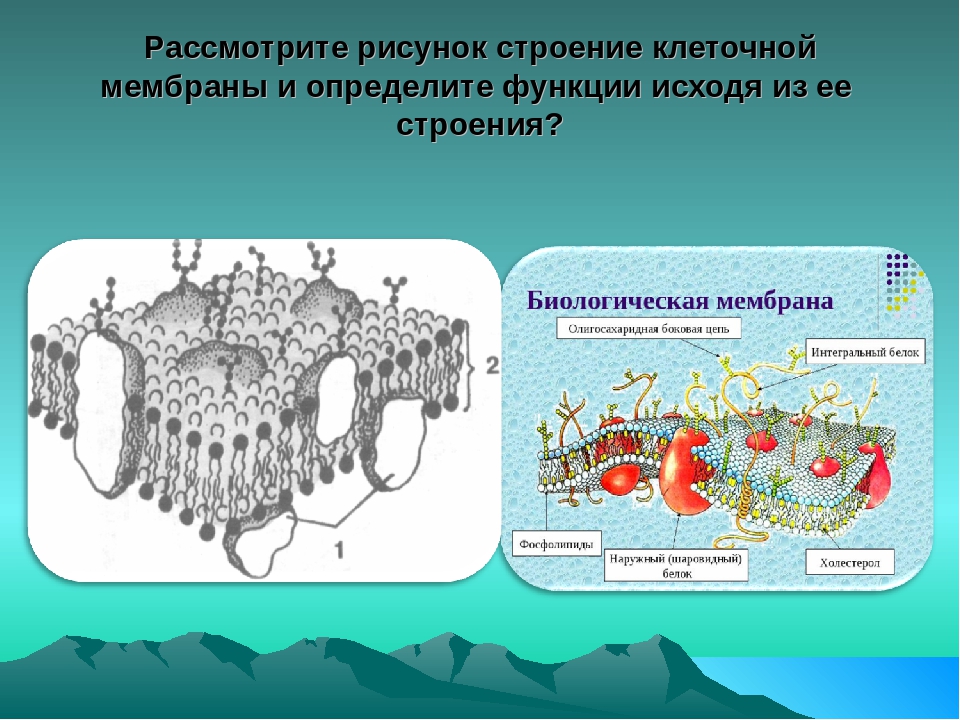
Выбор оборудования
Для битумосодержащих или мембранных крыш, по поверхности которых кабель укладывается напрямую, без теплорассеивающей подкладки, линейная мощность резистивного кабеля не должна превышать 20 Вт/м. Если нагревательный кабель устанавливается на металлическую полосу или лист, или на металлическую сварную сетку, то можно выбрать саморегулируемый или резистивный кабель с линейной мощностью до 30 Вт/м. Компания DEVI выпускает защищённые от воздействия ультрафиолетового излучения Солнца двухжильные резистивные кабели DEVIsnow™ 20T (20 Вт/м), DEVIsnow™ 30T (30 Вт/м), DEVIsafe™ 20T (20 Вт/м). Кабели удовлетворяют жёстким требованиям класса М2 международного стандарта 60800:2009 и имеют гарантийный срок 20 лет. Саморегулируемый кабель DEVIiceguard™ 18 также имеет оболочку, стойкую к УФ-излучению. Линейная мощность: 36 Вт/м при 0°С в талом снегу.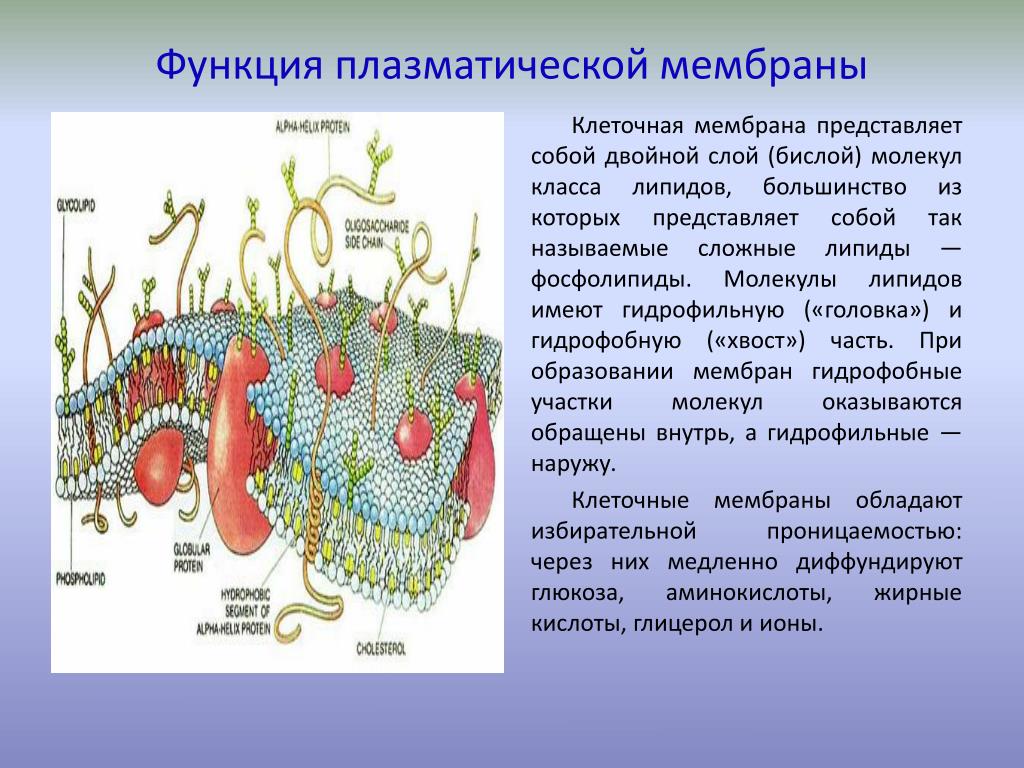 Гарантийный срок: 5 лет.
Гарантийный срок: 5 лет.
Приёмы монтажа АОС
Специалисты компании DEVI разработали специальные аксессуары в виде пластиковых клипс, зажимов и струбцин, а также монтажные ленты, предназначенные для закрепления нагревательных кабелей всех типов на участках рассматриваемых типах кровельных покрытий без нарушения их целостности. Имеются в виду настенные желоба, карнизы, капельники (где растут сосульки) и ендовы. При установке в водосточной системе наиболее часто кабели закрепляются в 2 или 4 линии на двойной монтажной ленте, которая в водосливах выполняет также роль разгрузочного элемента, заменяя трос, лавсановый канат или цепь. Ширина обогреваемых дорожек в ендовах и на карнизах зависит от многих факторов и может быть от 0,2 м до 1 м. Схема установки кабеля: параллельные линии или зигзаг. В любом случае шаг укладки кабеля должен выбираться таким, чтобы была обеспечена требуемая удельная установленная мощность, максимальное значение которой составляет 350 Вт/м². Для крепления кабеля в зоне обогреваемых дорожек применяется одинарная монтажная лента, алюминиевая клейкая лента повышенной прочности (на пологих участках кровли), двойная монтажная лента (на мягких и мембранных кровлях), специальные пластиковые зажимы для укладки нагревательных кабелей зигзагом (см. рисунки).
Для крепления кабеля в зоне обогреваемых дорожек применяется одинарная монтажная лента, алюминиевая клейкая лента повышенной прочности (на пологих участках кровли), двойная монтажная лента (на мягких и мембранных кровлях), специальные пластиковые зажимы для укладки нагревательных кабелей зигзагом (см. рисунки).
СТРУКТУРНЫЕ ОСОБЕННОСТИ МЕМБРАНЫ ЭРИТРОЦИТОВ ПРИ ПОСТПЕРФУЗИОННОМ ГЕМОЛИЗЕ РАЗЛИЧНОЙ СТЕПЕНИ ВЫРАЖЕННОСТИ
Чумакова С.П., Уразова О.И., Шипулин В.М., Новицкий В.В., Мальцева И.В., Вагина А.Ю., Никищенко С.А., Сапрыкина Э.В.
Общая патология и патологическая физиология
УДК 661.155
ГОУ ВПО Сибирский государственный медицинский университет Росздрава, г. Томск, Россия
Резюме:
В исследование вошли больные ишемической болезнью сердца с умеренным (21 человек) и выраженным (9 человек) гемолизом после коронарного шунтирования в условиях искусственного кровообращения. До и после операции изучали концентрацию свободного гемоглобина в плазме крови, содержание холестерина и фосфолипидов в липидных экстрактах и сорбционную емкость мембран эритроцитов, уровень ретикулоцитов в крови. Установлено, что у больных с выраженным постперфузионным гемолизом до операции содержание холестерина в мембране эритроцитов меньше, а фосфолипидов – больше, чем при умеренной гемоглобинемии. Это является не причиной эритролизиса, а следствием поступления в кровь молодых форм эритроцитов в связи с ускоренной гибелью до операции клеток, обладающих высокой проницаемостью мембраны (в отличие от группы с умеренным гемолизом). Под влиянием искусственного кровообращения проницаемость мембраны еще больше увеличивается, вызывая развитие выраженного внутрисосудистого гемолиза после операции.
До и после операции изучали концентрацию свободного гемоглобина в плазме крови, содержание холестерина и фосфолипидов в липидных экстрактах и сорбционную емкость мембран эритроцитов, уровень ретикулоцитов в крови. Установлено, что у больных с выраженным постперфузионным гемолизом до операции содержание холестерина в мембране эритроцитов меньше, а фосфолипидов – больше, чем при умеренной гемоглобинемии. Это является не причиной эритролизиса, а следствием поступления в кровь молодых форм эритроцитов в связи с ускоренной гибелью до операции клеток, обладающих высокой проницаемостью мембраны (в отличие от группы с умеренным гемолизом). Под влиянием искусственного кровообращения проницаемость мембраны еще больше увеличивается, вызывая развитие выраженного внутрисосудистого гемолиза после операции.
Ключевые слова:
гемолиз, эритроциты, мембрана, искусственное кровообращение.
Авторская справка
Чумакова С. П.
П.
Уразова Ольга Ивановна
Шипулин Владимир Митрофанович
Новицкий Вячеслав Викторович
Мальцева Инна Викторовна
Вагина Алена Юрьевна
Никищенко Светлана Александровна
Сапрыкина Элеонора Васильевна
ГОУ ВПО Сибирский государственный медицинский университет Росздрава
Россия, 634061, г. Томск, ул. Тверская д.53, кв.40
E-mail: [email protected]
Секреты производства мембраны LOGICROOF и теплоизоляции LOGICPIR — ТЕХНОНИКОЛЬ
Благодаря современным технологиям производства материалы компании ТЕХНОНИКОЛЬ легко противостоят всем капризам природы, эффективно защищают дома и сохраняют максимальный комфорт в помещении. Как этого удается достичь?
Рассказываем о том, как мы производим мембраны LOGICROOF и теплоизоляцию LOGICPIR, чтобы они максимально эффективно выполняли свои функции.
В чем особенность мембраны LOGICROOF
Эта высокотехнологичная мембрана прекрасно работает в паре с утеплителем LOGICPIR. У нее самая высокая скорость монтажа среди гидроизоляционных материалов, а устанавливать ее можно в любой сезон и при любых погодных условиях. При этом есть широкий ассортимент решений для всех типов систем.
Как мы производим материал
Заводы, на которых изготавливают мембраны LOGICROOF и теплоизоляцию LOGICPIR, находятся по соседству друга от друга в Рязани. Это передовые производства с самой скоростной линией в России и Европе. Мощность – 30 млн м² материала в год. Здесь трудится штат высококлассных специалистов и есть собственная научная лаборатория.
Все компоненты для изготовления мембраны хранят отдельно. Оператор только выбирает рецепт их смешивания, а затем весь процесс происходит в автоматическом режиме. Программа строго соблюдает пропорции благодаря системе высокоточной дозации.
Пока готовится смесь для будущей полимерной мембраны, в производственную линию заправляют армирующую сетку. Именно на нее будет нанесен расправленный ПВХ-порошок.
Именно на нее будет нанесен расправленный ПВХ-порошок.
Важно выдерживать правильную температуру, ведь ПВХ не любит перегрева и сильного трения. Технологии ТЕХНОНИКОЛЬ позволяют избежать повреждения мембраны: например, оба экструдера на заводе имеют контрвращение, благодаря которому частицы в полимерной матрице распределяются равномернее.
Расплавленный полимер накладывается на основу будущей мембраны через специальную формующую фильеру – так называют высокопрочные формы с тарированными отверстиями, расположенными в определенном порядке.
Ее задача – выдерживать установленную ширину полотна и заставить вязкую субстанцию заполнить все ячейки армированной сетки.
Скелет сетки запаивают между двумя и более слоями, чтобы достичь главных характеристик LOGICROOF: прочности и гибкости. За толщиной полученных слоев следят два специальных изотопных толщиномера. Данные из них поступают на монитор оператора.
Готовую мембрану охлаждают, обрезают в нужную ширину, маркируют и наматывают в большие рулоны – биг-роллы. Здесь же из партии берут образец и отправляют его на исследование в лабораторию.
Здесь же из партии берут образец и отправляют его на исследование в лабораторию.
Биг-роллы снимают с производственной линии и перемещают на установку перемотки и упаковки.
В чем особенность теплоизоляции LOGICPIR
Это теплоизоляция нового поколения. Для качественного утепления любого здания хватает гораздо меньшего слоя этого материала по сравнению с другими решениями.
При этом LOGICPIR не поддерживает горение и не распространяет пламя. Он даже не выделяет вредных веществ при нагревании, что подтверждено международным сертификатом Eurofins Expert Services (Финляндия) и заключением «НМИЦ здоровья детей».
Как мы производим материал
В составе утеплителя – жидкая смесь из 16 компонентов. Сочетая их по-разному, можно создать уникальный, подходящий именно под ваши цели продукт.
Более того, ТЕХНОНИКОЛЬ уделяет особое внимание удобству своих покупателей, поэтому использует фольгу с антибликовым покрытием. Благодаря этому с материалом легко работать в солнечную погоду.
Благодаря этому с материалом легко работать в солнечную погоду.
Производство полностью автоматизировано, что практически исключает ошибки на линии.
Из динамического миксера смесь через клапаны выливают на фольгированную обкладку. Она становится облицовкой утеплителя, защищающей его от пара и влаги.
Реакционная смесь пенится и растет между обкладками. Посмотреть на это можно лишь в лаборатории: в стакане на изображении ниже смесь ведет себя так же, как и на линии.
Как будет расти материал, зависит от подобранной рецептуры. Она всегда учитывает параметры реакции, при которой увеличивается объем пены.
Как только начинается активный процесс, перед входом в ламинатор вместе с пеной заправляется облицовка. Двойной ленточный ламинатор с точностью до 1 мм позволяет сформировать толщину плиты.
Здесь при температуре 70 °C формируются миллионы маленьких ячеек с газом, благодаря чему материал достигает уникальной теплопроводности в 0,022 Вт/м*К.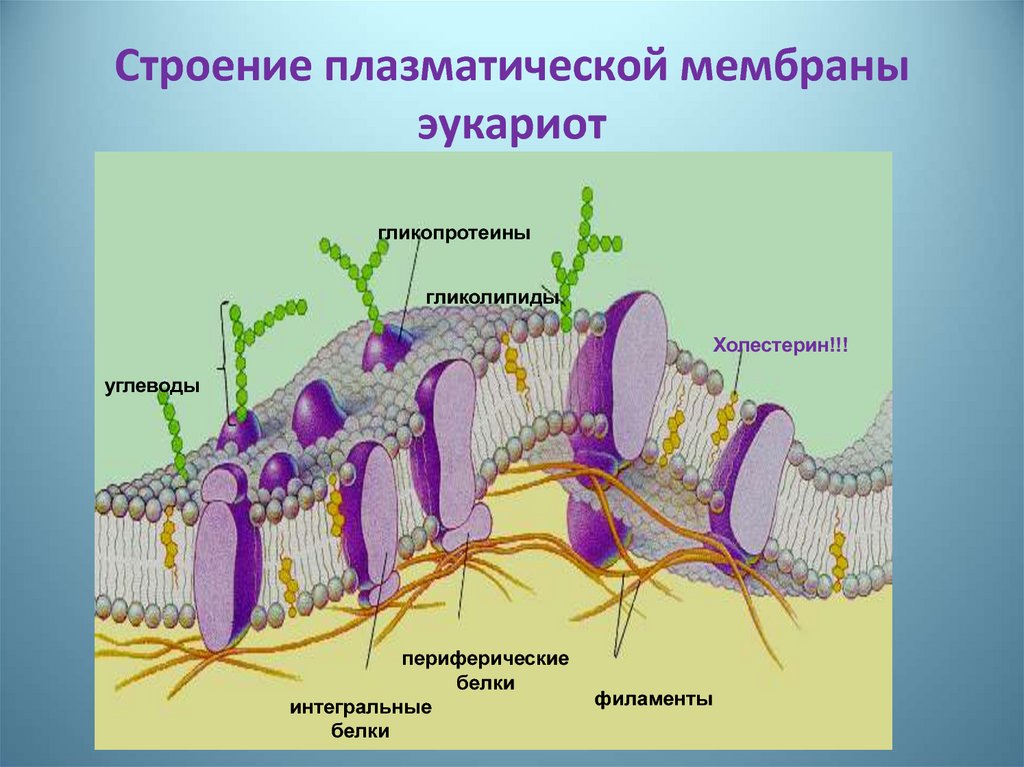 После выхода из двойного ленточного ламинатора его режут на так называемые mother-борды, то есть плиты длиной до 6 метров.
После выхода из двойного ленточного ламинатора его режут на так называемые mother-борды, то есть плиты длиной до 6 метров.
От одной из плит отрезают образцы и отправляют на испытания. Сотрудники лаборатории должны подтвердить заявленные характеристики и гарантировать надежность всей партии.
Причем берут сразу несколько равных кусков: из левой, срединной и правой частей. Полученный материал помещают в специальную динамометрическую машину, работающую на сжатие. Она проверяет материал на прочность.
Чтобы контролировать изменения геометрии утеплителя, образцы из всех партий перемещают в холодильник с температурой −40 °C. Материал очень легкий, так что любой человек может спокойно нести его одной рукой.
Утеплитель не должен потерять за время испытания более 1 % от первоначальных размеров. Нахождение в таких специально созданных условиях позволяет понять, как продукт будет вести себя зимой или при утеплении кровли.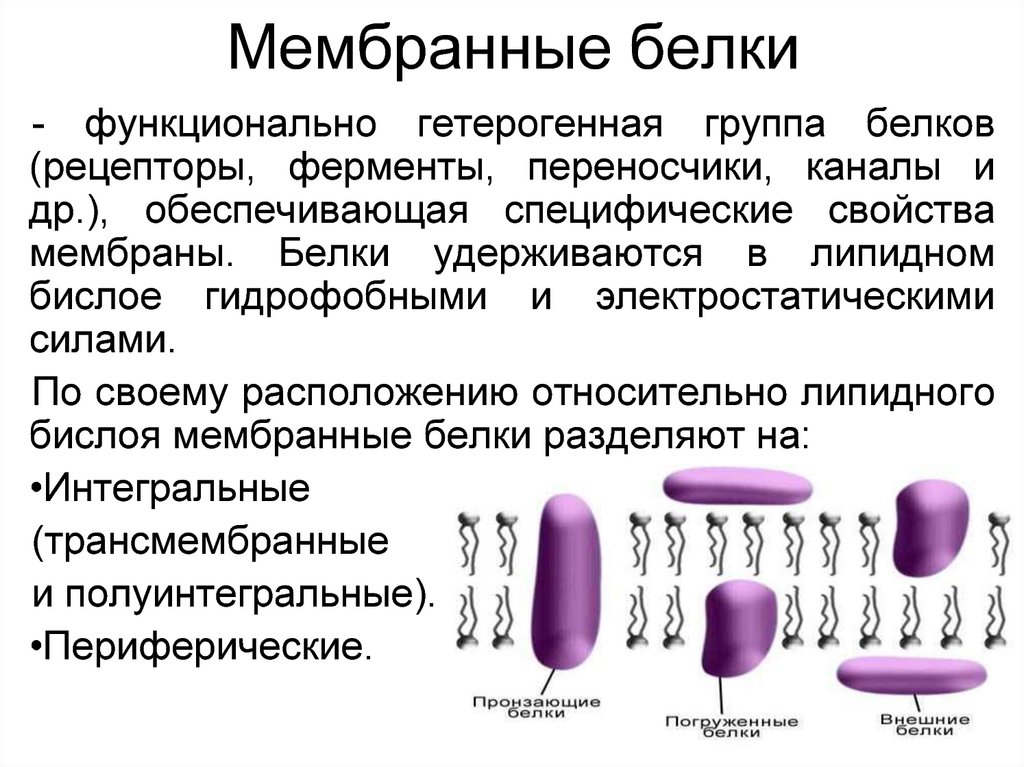
Также утеплитель LOGICPIR испытывают в огневой камере. Под воздействием пламени поверхность плиты обугливается, незначительно вспучивается и дальше перестает реагировать на жар. Насквозь она не прогорает. Температура может достигать здесь 1000 °С, однако слой материала долго выдерживает даже эти условия.
Мембрана LOGICROOF и утеплитель LOGICPIR – это надежные материалы, которые сделают ваше строение долговечным и максимально комфортным для жизни. Переходите на сайт ТЕХНОНИКОЛЬ, чтобы узнать больше об этих и многих других инновационных технологиях компании.
Membrane Fluidity | Protocol (Translated to Russian)
5.2: Текучесть мембран
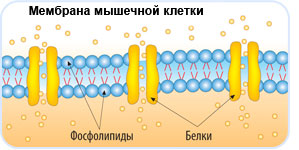
Клеточные мембраны состоят из фосфолипидов, белков и углеводов, которые слабо связаны друг с другом посредством химических взаимодействий. Молекулы обычно могут перемещаться в плоскости мембраны, придавая мембране ее гибкий характер, называемый текучестью.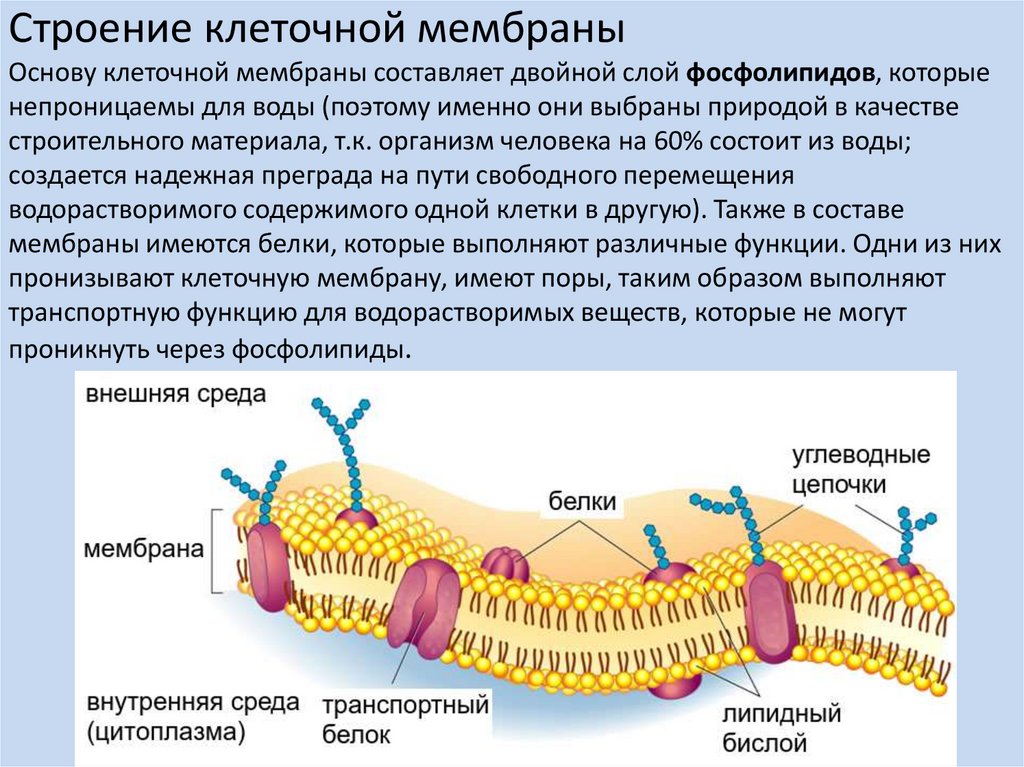 Две другие особенности мембраны способствуют текучести мембраны: химическая структура фосфолипидов и присутствие холестерина в мембране.
Две другие особенности мембраны способствуют текучести мембраны: химическая структура фосфолипидов и присутствие холестерина в мембране.
Жирные кислоты фосфолипидов могут быть насыщенными или ненасыщенными. Насыщенные жирные кислоты имеют одинарные связи между углеводородным остовом и насыщены максимальным количеством атомов водорода. Эти насыщенные хвосты прямые и поэтому могут плотно сбиваться. Напротив, хвосты ненасыщенных жирных кислот содержат двойные связи между атомами углерода, что придает им изогнутую форму и предотвращает плотную упаковку. Увеличение относительной доли фосфолипидов с ненасыщенными хвостами приводит к более жидкой мембране. Организмы, такие как бактерии и дрожжи, которые испытывают колебания температуры окружающей среды, могут регулировать содержание жирных кислот в своих мембранах, чтобы поддерживать относительно постоянную текучесть.
В клеточных мембранах холестерин может взаимодействовать с головками фосфолипидов, частично иммобилизуя проксимальную часть углеводородной цепи. Это взаимодействие снижает способность полярных молекул пересекать мембрану. Холестерин также предотвращает плотную упаковку фосфолипидов, тем самым предотвращая вероятность замерзания мембран. Точно так же холестерин действует как структурный буфер при повышении температуры, ограничивая чрезмерную текучесть.
Это взаимодействие снижает способность полярных молекул пересекать мембрану. Холестерин также предотвращает плотную упаковку фосфолипидов, тем самым предотвращая вероятность замерзания мембран. Точно так же холестерин действует как структурный буфер при повышении температуры, ограничивая чрезмерную текучесть.
Также предполагается, что холестерин играет роль в организации мембранных липидов и белков в функциональные группы, называемые липидными рафтами. Считается, что эти группы белков, фосфолипидов и холестерина разделяют области мембраны, располагая молекулы с аналогичными функциями в непосредственной близости друг от друга. Однако конкретная структура и функция этих мембранных пластырей неясны и являются активной областью исследований.
Литература для дополнительного чтения
Renne, Mike F., and Anton IPM de Kroon. “The role of phospholipid molecular species in determining the physical properties of yeast membranes. ” FEBS Letters 592, no. 8 (2018): 1330-1345. [Source]
” FEBS Letters 592, no. 8 (2018): 1330-1345. [Source]
Steck, Theodore L., and Yvonne Lange. “Cell cholesterol homeostasis: mediation by active cholesterol.” Trends in Cell Biology 20, no. 11 (2010): 680-687. [Source]
| Влияние поливинилпирролидона на морфологию, гидрофильность и эксплуатационные характеристики микрофильтрационных мембран из полиэфирсульфона Е. Ю. Астахов, И. М. Колганов, Э. Р. Клиншпонт, П. Г. Царин, А. А. Калачева |
163 |
| Особенности электродиализа растворов винной кислоты и ее солей Т. В. Елисеева, Е. В. Крисилова, В. П. Василевский, Э. Г. Новицкий |
173 |
| Влияние условий получения на наноструктурные особенности перфторированных протонопроводящих мембран типа Nafion® Ю. В. Кулъвелис, В. Т. Лебедев, В. А. Трунов, О. Н. Примаченко, С. Я. Хайкин, Д. Торок, С.  С. Иванчев С. Иванчев
|
179 |
| Диффузионные характеристики поверхностно-модифицированных ионообменных мембран на основе МК-40, МФ-4СК и полианилина П. А. Юрова, Ю. А. Караванова, А. Б. Ярославцев |
186 |
| Мозаичный характер гелевой поляризации при ультрафильтрации А. Н. Черкасов, А. Е. Полоцкий Целью настоящего исследования является уточнение поляризационно-ситовой модели ультрафильтрации (УФ), предложенной ранее одним из авторов. Согласно этой модели, гель, покрывающий сплошным слоем поверхность мембраны в процессе ультрафильтрации, уменьшает радиус всех пор мембраны на постоянную величину Δ R, что объясняет, в частности, сохранение ситового механизма УФ при гелевой поляризации. Для проверки справедливости предположения о постоянстве величины Δ R проведен расчет кривых задержания (КЗ) для мембран с разной шириной распределения пор по размерам при разном уровне гелевой поляризации. |
191 |
| Фракционирование многокомпонентных растворов методом электродиализа с биполярными мембранами Е. Н. Егоров, А. А. Свитцов, С.  Н. Дудник, В. И. Демкин Н. Дудник, В. И. Демкин
|
198 |
| Нанофильтрация и сорбция органических растворителей в образцах поли-1-триметилсилил-1-пропина различной микроструктуры А. В. Волков, С. Е. Царьков, М. Б. Гокжаев, Г. Н. Бондаренко, С. А. Легкое, Ю. А. Кукушкина, В. В. Волков Исследовано сорбционное и нанофильтрационное поведение двух образцов ПТМСП, синтезированных с использованием различных каталитических систем (TaCl5/TIBA и NbCl5) и различающихся микроструктурой цепи (соотношение cis/trans составляет 50/50 и 63/37 для ПТМСП/Ta и ПТМСП/Nb, соответственно). Измерены величины равновесной сорбции (гомологический ряд линейных спиртов С1 ‾ С10, диолы и кетоны для ПТМСП/Ta и линейные спирты С1 ‾ С10 для ПТМСП/Nb) и величины набухания полимеров в этих средах. Исследована нанофильтрация разбавленных растворов трех красителей (Solvent Blue 35, Safranine O и Remazol Brilliant Blue R) в этаноле для образца ПТМСП/Ta и проведено сравнение нанофильтрационных характеристик с аналогичными данными, полученными ранее для ПТМСП/Nb. |
209 |
| Применение электрохимической импедансной спектроскопии для определения толщины диффузионного слоя около поверхности ионообменных мембран А. Э. Козмай, В. В. Никоненко, Н. Д. Писъменская, С. А. Мареев, Е. И. Белова, P. Sistat Методом электрохимической импедансной спектроскопии изучены три ионообменные мембраны: гомогенная анионообменная мембрана АМХ, гетерогенная катионообменная мембрана МК-40 и гомогенная катионообменная мембрана Nafion-117. Обработка экспериментальных спектров импеданса с помощью разработанной ранее модели позволила найти нернстовскую толщину диффузионного пограничного слоя δ (ДПС) как функцию плотности тока. |
221 |
| Влияние состава раствора на селективность обратноосмотических и нанофильтрационных мембран Г. Г. Каграманов, Е. Н. Фарносова |
233 |
| Правила по подготовке рукописи в журнале “Мембраны и мембранные технологии” | 240 |
 Обнаружены немонотонная зависимость сдвига КЗ, а также изменение их ширины в ходе поляризации, что противоречит экспериментальным данным. Сделано предположение, что уменьшение размера поры при гелевой поляризации (ГП) происходит на величину, пропорциональную ее размеру (Δ R ~ R). Расчет кривых задержания в этом предположении показал параллельный монотонный сдвиг КЗ с ростом поляризации с сохранением их ширины, что подтверждается экспериментом. Полученные данные свидетельствуют в пользу мозаичного характера гелевой поляризации, когда гель покрывает поверхность мембраны не сплошным однородным слоем, как это считается в классической теории ГП, а образуется преимущественно у входа в пору и его локальный неоднородный характер определяется размерами конкретной поры, а не усредненными параметрами мембраны.
Обнаружены немонотонная зависимость сдвига КЗ, а также изменение их ширины в ходе поляризации, что противоречит экспериментальным данным. Сделано предположение, что уменьшение размера поры при гелевой поляризации (ГП) происходит на величину, пропорциональную ее размеру (Δ R ~ R). Расчет кривых задержания в этом предположении показал параллельный монотонный сдвиг КЗ с ростом поляризации с сохранением их ширины, что подтверждается экспериментом. Полученные данные свидетельствуют в пользу мозаичного характера гелевой поляризации, когда гель покрывает поверхность мембраны не сплошным однородным слоем, как это считается в классической теории ГП, а образуется преимущественно у входа в пору и его локальный неоднородный характер определяется размерами конкретной поры, а не усредненными параметрами мембраны. Анализ результатов по сорбции и набуханию ПТМСП/Ta позволил сделать предположение о наличии сорбционных центров в ПТМСП, способных координировать около себя две гидроксильные группы. Методами колебательной ИК-спектроскопии и квантовой химии исследована структура ПТМСП/Ta и ПТМСП/Nb. показано, что природа центров сорбции в ПТМСП, скорее всего, связана с необычайно высокой полярностью связей С=С и SiC в элементарном звене полимера. Расчет модели из 5 звеньев ПТМСП в присутствии мономера этилового спирта показал, что в такой системе реализуется комплекс с Δ E = 18 кДж/моль. Результаты нанофильтрационных исследований свидетельствуют, что коэффициент проницаемости этанола и общий поток растворов красителей в этаноле приблизительно в два раза ниже для ПТМСП/Ta по сравнению с ПТМСП/Nb. Для обоих образцов ПТМСП удерживание красителей возрастает в ряду Solvent Blue 35, Safranine O и Remazol Brilliant Blue R в соответствии с увеличением коэффициентов распределения красителей. Удерживание красителей Solvent Blue 35 и Safranine O с молекулярными массами 350 г/моль заметно ниже для ПТМСП/Ta по сравнению с ПТМСП/Nb.
Анализ результатов по сорбции и набуханию ПТМСП/Ta позволил сделать предположение о наличии сорбционных центров в ПТМСП, способных координировать около себя две гидроксильные группы. Методами колебательной ИК-спектроскопии и квантовой химии исследована структура ПТМСП/Ta и ПТМСП/Nb. показано, что природа центров сорбции в ПТМСП, скорее всего, связана с необычайно высокой полярностью связей С=С и SiC в элементарном звене полимера. Расчет модели из 5 звеньев ПТМСП в присутствии мономера этилового спирта показал, что в такой системе реализуется комплекс с Δ E = 18 кДж/моль. Результаты нанофильтрационных исследований свидетельствуют, что коэффициент проницаемости этанола и общий поток растворов красителей в этаноле приблизительно в два раза ниже для ПТМСП/Ta по сравнению с ПТМСП/Nb. Для обоих образцов ПТМСП удерживание красителей возрастает в ряду Solvent Blue 35, Safranine O и Remazol Brilliant Blue R в соответствии с увеличением коэффициентов распределения красителей. Удерживание красителей Solvent Blue 35 и Safranine O с молекулярными массами 350 г/моль заметно ниже для ПТМСП/Ta по сравнению с ПТМСП/Nb. Найденные закономерности сорбционного и нанофильтрационного поведения ПТМСП/Ta и ПТМСП/Nb находят логичное объяснение в рамках существующих подходов, рассматривающих аморфные стеклообразные полимеры как микрогетерогенные среды с областями различной степени упорядоченности полимерных цепей, а наблюдаемые различия их сорбционных и мембранных свойств определяются более рыхлой упаковкой полимерных цепей образца ПТМСП/Ta по сравнению с образцом ПТМСП/Nb.
Найденные закономерности сорбционного и нанофильтрационного поведения ПТМСП/Ta и ПТМСП/Nb находят логичное объяснение в рамках существующих подходов, рассматривающих аморфные стеклообразные полимеры как микрогетерогенные среды с областями различной степени упорядоченности полимерных цепей, а наблюдаемые различия их сорбционных и мембранных свойств определяются более рыхлой упаковкой полимерных цепей образца ПТМСП/Ta по сравнению с образцом ПТМСП/Nb. Показано, что поведение мембраны АМХ близко к “идеальному”, описываемому в рамках модели: спектр импеданса мембраны близок к теоретическому, а величина δ лишь ненамного меньше величины δ Lev, рассчитанной с помощью уравнения Левека, полученного в рамках классической теории конвективной диффузии. Поведение МК-40 и Nafion заметно отличается от “идеального”: реактивная составляющая импеданса заметно меньше теоретической в области средних частот, отвечающих точке максимума на низкочастотной части спектра вида спектра Варбурга для отрезка конечной длины. Величина δ < δ Lev даже при допредельных токах, и это отклонение увеличивается с ростом плотности тока. Эти особенности поведения мембран хорошо коррелируют с результатами вольтамперометрии. Поведение изученных мембран связано со свойствами их поверхности: гетерогенность (случай МК-40) и, в особенности, высокая гидрофобность поверхности (Nafion-117) способствуют развитию электроконвекции. Высокая гидрофильность поверхности мембраны АМХ обусловливают ее поведение, близкое к “идеальному”.
Показано, что поведение мембраны АМХ близко к “идеальному”, описываемому в рамках модели: спектр импеданса мембраны близок к теоретическому, а величина δ лишь ненамного меньше величины δ Lev, рассчитанной с помощью уравнения Левека, полученного в рамках классической теории конвективной диффузии. Поведение МК-40 и Nafion заметно отличается от “идеального”: реактивная составляющая импеданса заметно меньше теоретической в области средних частот, отвечающих точке максимума на низкочастотной части спектра вида спектра Варбурга для отрезка конечной длины. Величина δ < δ Lev даже при допредельных токах, и это отклонение увеличивается с ростом плотности тока. Эти особенности поведения мембран хорошо коррелируют с результатами вольтамперометрии. Поведение изученных мембран связано со свойствами их поверхности: гетерогенность (случай МК-40) и, в особенности, высокая гидрофобность поверхности (Nafion-117) способствуют развитию электроконвекции. Высокая гидрофильность поверхности мембраны АМХ обусловливают ее поведение, близкое к “идеальному”.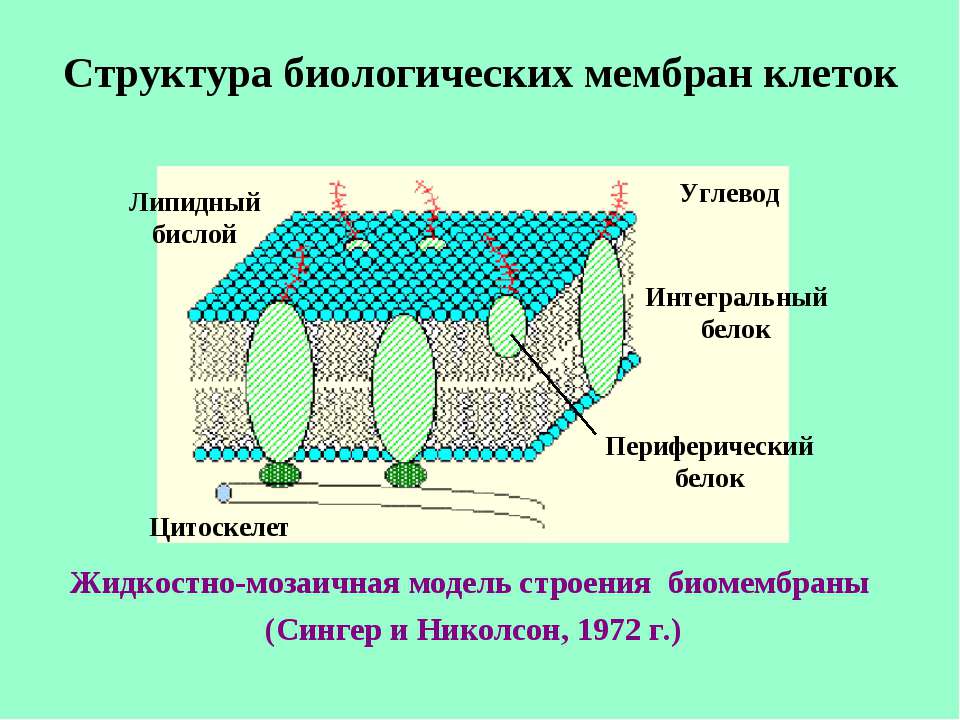
4: Мембраны – структура, свойства и функции
- Последнее обновление
- Сохранить как PDF
Биологические мембраны являются основой многих важных свойств клетки, не последним из которых является физическое определение границ клетки, а у эукариот – границ каждой внутриклеточной органеллы.Однако они не являются полностью непроницаемыми границами, и через встроенные белки мембрана служит привратником для прохождения определенных молекул в (например, питательные вещества) и из (например, отходы) клетки. Другие встроенные белки могут идентифицировать клетку для других клеток и участвовать во множестве взаимодействий с окружающей средой или другими клетками. Наконец, мембрана, или, точнее, химические градиенты через мембрану, является важным источником энергии для клетки.
Другие встроенные белки могут идентифицировать клетку для других клеток и участвовать во множестве взаимодействий с окружающей средой или другими клетками. Наконец, мембрана, или, точнее, химические градиенты через мембрану, является важным источником энергии для клетки.
- 4.1: Структура и состав мембраны
- Поскольку большинство клеток живут в водной среде, и содержимое клетки также в основном водное, очевидно, что мембрана, отделяющая одну сторону от другой, должна быть гидрофобной, чтобы сформировать эффективный барьер против случайная утечка материалов или воды. Клеточные мембраны были частично определены как состоящие в основном из фосфолипидов: молекул, состоящих из фосфорилированной полярной головной группы, присоединенной к глицериновой основной цепи, имеющей два длинных углеводородных хвоста.
- 4.2: Проницаемость мембраны
- Чистый фосфолипидный бислой, независимо от липидного состава, представляет собой полупроницаемую мембрану, которая обычно отталкивает большие молекулы и ионы.
 Небольшие полярные молекулы иногда могут проходить легко (например, этанол), но чаще проходят с низкой скоростью (например, вода). Однако небольшие неполярные молекулы могут относительно легко проходить через мембрану. Причины должны быть очевидны: более крупные молекулы просто не могут поместиться между молекулами липидов, чтобы пробиться сквозь них.
Небольшие полярные молекулы иногда могут проходить легко (например, этанол), но чаще проходят с низкой скоростью (например, вода). Однако небольшие неполярные молекулы могут относительно легко проходить через мембрану. Причины должны быть очевидны: более крупные молекулы просто не могут поместиться между молекулами липидов, чтобы пробиться сквозь них.
- 4.3: Мембранные транспортные белки
- Мембранные белки бывают двух основных типов: интегральные мембранные белки (иногда называемые внутренними), которые непосредственно встроены в фосфолипидный бислой, и периферические мембранные белки (иногда называемые внешними), которые расположены очень близко или даже в контакте с одной стороной мембраны, но не проникают в гидрофобное ядро бислоя. Интегральные мембранные белки могут полностью проходить через мембрану, контактируя как с внеклеточной средой, так и с цитоплазмой.
- 4.4: Потенциал действия в нейронах
- Транспортировка растворенных веществ в клетки и из них имеет решающее значение для жизни.
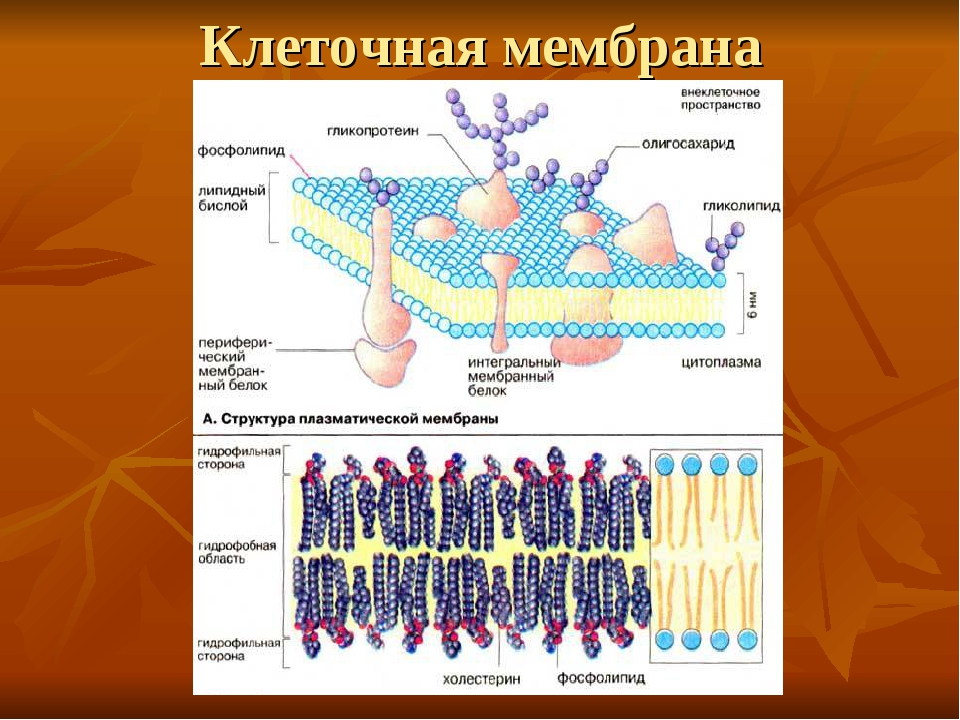 Однако в нейронах движение ионов выполняет еще одну важную функцию у многоклеточных животных: производство потенциалов действия, используемых для нейротрансмиссии. Эта специализация позволяет чрезвычайно быстро передавать информацию на большие расстояния. Примером, который мой наставник использовал при обучении школьников основам неврологии, был биполярный нейрон, который простирается от пальца ноги до мозга.
Однако в нейронах движение ионов выполняет еще одну важную функцию у многоклеточных животных: производство потенциалов действия, используемых для нейротрансмиссии. Эта специализация позволяет чрезвычайно быстро передавать информацию на большие расстояния. Примером, который мой наставник использовал при обучении школьников основам неврологии, был биполярный нейрон, который простирается от пальца ноги до мозга.
Миниатюра: клеточная мембрана, также называемая плазматической мембраной или плазмалеммой, представляет собой полупроницаемый липидный бислой, общий для всех живых клеток. Он содержит множество биологических молекул, в первую очередь белков и липидов, которые участвуют во множестве клеточных процессов. Он также служит точкой прикрепления как внутриклеточного цитоскелета, так и, если имеется, клеточной стенки. (Общественное достояние; LadyofHats через Википедию.
Модель прогноза для мембранных белков с использованием функций, основанных на моментах
Самой подходящей единицей человеческого тела является его клетка.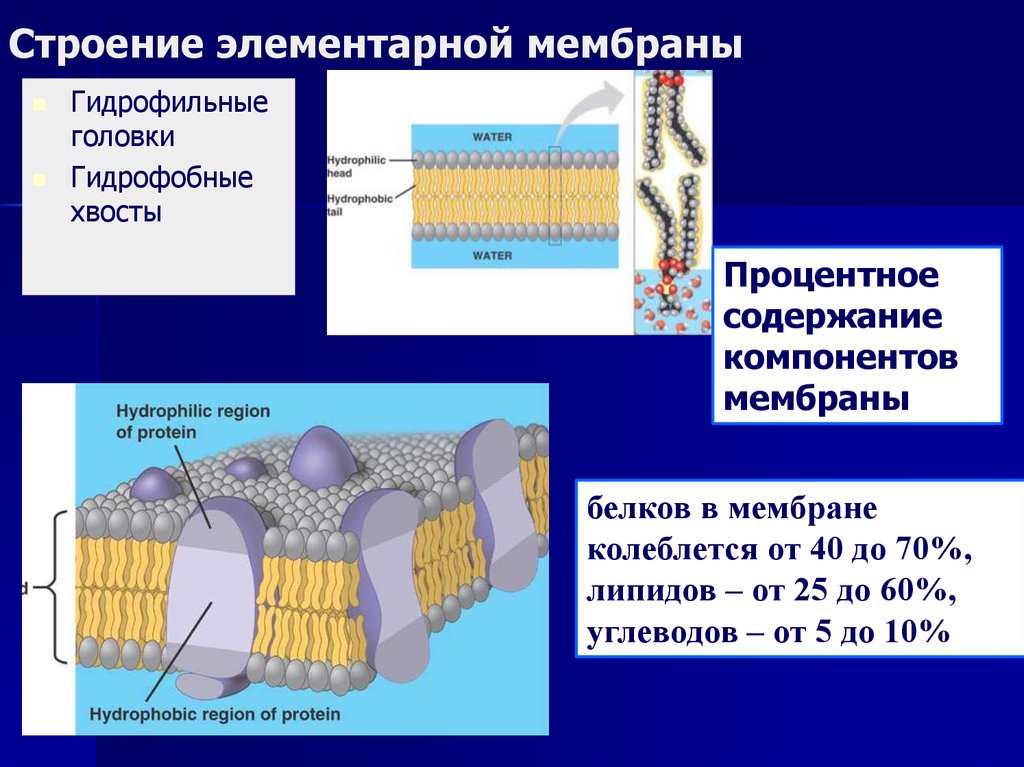 Внутри клетки заключено множество бесконечно малых объектов и молекул, которые защищены клеточной мембраной. Белки, которые связаны с этой двухслойной клеточной мембраной на основе липидов, известны как мембранные белки и, как считается, играют важную роль. Эти мембранные белки проявляют свое влияние на клеточную активность внутри и вне клетки. По словам ученых из фармацевтических организаций, эти мембранные белки выполняют ключевую задачу во взаимодействии с лекарствами. В этом исследовании представлена методика, основанная на различных вычислительно интеллектуальных методах, используемых для предсказания мембранного белка без экспериментального использования масс-спектрометрии.Статистические моменты использовались для извлечения признаков, и, кроме того, многослойная нейронная сеть была обучена с использованием обратного распространения ошибки для предсказания мембранных белков. Результаты показывают, что предлагаемый метод работает лучше, чем существующие методики.
Внутри клетки заключено множество бесконечно малых объектов и молекул, которые защищены клеточной мембраной. Белки, которые связаны с этой двухслойной клеточной мембраной на основе липидов, известны как мембранные белки и, как считается, играют важную роль. Эти мембранные белки проявляют свое влияние на клеточную активность внутри и вне клетки. По словам ученых из фармацевтических организаций, эти мембранные белки выполняют ключевую задачу во взаимодействии с лекарствами. В этом исследовании представлена методика, основанная на различных вычислительно интеллектуальных методах, используемых для предсказания мембранного белка без экспериментального использования масс-спектрометрии.Статистические моменты использовались для извлечения признаков, и, кроме того, многослойная нейронная сеть была обучена с использованием обратного распространения ошибки для предсказания мембранных белков. Результаты показывают, что предлагаемый метод работает лучше, чем существующие методики.
1. Введение
Среди различных макромолекул, составляющих клетку, белки являются структурной и функциональной единицей клетки. Белки проводят тысячи химических реакций и обрабатываются как внутри, так и вне клетки.Каждая клетка окружена защитной стенкой, состоящей из липидов и названной плазматической мембраной. Тем не менее, очень немногие липидорастворимые и неполярные молекулы могут проникать путем прямой диффузии через липидный бислой. В большинстве случаев перенос молекул через мембраны осуществляется мембранными белками [1]. Примерно 25–75% массы мембраны состоит из белков. Эти белки могут быть интегральными или периферическими. Эти белки могут действовать как рецепторы и играть важную роль в передаче сигналов клетками.Гликопротеины отвечают за клеточную адгезию. Некоторые белки играют важную роль в транспортировке молекул через клеточные мембраны. Эти белки могут быть носителями, каналами или насосами. Около 60–70% этих белков состоят из α спиралей; очень немногие состоят из β стволов [2].
Белки проводят тысячи химических реакций и обрабатываются как внутри, так и вне клетки.Каждая клетка окружена защитной стенкой, состоящей из липидов и названной плазматической мембраной. Тем не менее, очень немногие липидорастворимые и неполярные молекулы могут проникать путем прямой диффузии через липидный бислой. В большинстве случаев перенос молекул через мембраны осуществляется мембранными белками [1]. Примерно 25–75% массы мембраны состоит из белков. Эти белки могут быть интегральными или периферическими. Эти белки могут действовать как рецепторы и играть важную роль в передаче сигналов клетками.Гликопротеины отвечают за клеточную адгезию. Некоторые белки играют важную роль в транспортировке молекул через клеточные мембраны. Эти белки могут быть носителями, каналами или насосами. Около 60–70% этих белков состоят из α спиралей; очень немногие состоят из β стволов [2].
Аминокислоты являются структурным компонентом каждого белка. Структура белка строго связана с функцией этого белка. Если природа или последовательность белка изменяется, функция белка может измениться.В белках есть четыре типа структурных организаций: первичный, вторичный, третичный и четвертичный уровни структур [1]. Последовательность белка определяет конкретную функцию и форму структуры белка. Неформально белки можно разделить на три различных класса: глобулярные, волокнистые и мембранные белки. Эти типы белков связаны друг с другом, но имеют различия в своей третичной структуре [3–5]. Глобулярные белки – это в основном ферменты. Волокнистые белки часто являются структурными и в основном являются частью какой-либо клеточной структуры.Мембранные белки отвечают за передачу сигналов между клетками и действуют как каналы для полярных и неполярных молекул, которые транспортируются через клеточную мембрану [6, 7]. В частности, достижения в области молекулярной биологии привели к быстрому увеличению количества белковых последовательностей в банках данных. Эти белковые последовательности используются для извлечения различных свойств, связанных с этим белком.
Если природа или последовательность белка изменяется, функция белка может измениться.В белках есть четыре типа структурных организаций: первичный, вторичный, третичный и четвертичный уровни структур [1]. Последовательность белка определяет конкретную функцию и форму структуры белка. Неформально белки можно разделить на три различных класса: глобулярные, волокнистые и мембранные белки. Эти типы белков связаны друг с другом, но имеют различия в своей третичной структуре [3–5]. Глобулярные белки – это в основном ферменты. Волокнистые белки часто являются структурными и в основном являются частью какой-либо клеточной структуры.Мембранные белки отвечают за передачу сигналов между клетками и действуют как каналы для полярных и неполярных молекул, которые транспортируются через клеточную мембрану [6, 7]. В частности, достижения в области молекулярной биологии привели к быстрому увеличению количества белковых последовательностей в банках данных. Эти белковые последовательности используются для извлечения различных свойств, связанных с этим белком. Общее количество белковых последовательностей в банке данных Swiss-Prot составляло 3939 в 1986 году. В соответствии с версией 2015_03, выпущенной 4 марта 2015 года, общее количество белковых последовательностей достигло 547 964 записей.
Общее количество белковых последовательностей в банке данных Swiss-Prot составляло 3939 в 1986 году. В соответствии с версией 2015_03, выпущенной 4 марта 2015 года, общее количество белковых последовательностей достигло 547 964 записей.
Предсказание мембранного белка – сложная задача, и поэтому модель прогнозирования должна быть точной и эффективной. Большинство клеточных функций осуществляется через эти важные мембранные белки. Основной целью многих фармацевтических исследовательских организаций является мембранный белок, потому что 50% лекарств имеют своей мишенью мембранные белки [8–10]. Каждый тип мембранного белка имеет свое специфическое поведение и функциональность по отношению к клеточной мембране. Многие системы были предложены в [11–14] для определения точного назначения и поведения мембранного белка внутри мембраны и вне мембраны и дали результаты с точностью, но все еще есть много областей, где эту точность можно улучшить и получить эффективные результаты. может быть достигнуто.
Предлагаемая система пытается предсказать, соответствует ли данная последовательность белка мембранному или немембранному белку, давая точные и эффективные результаты по сравнению с существующими системами. Во-первых, из входного запроса белка извлекаются различные особенности, относящиеся к последовательности и содержанию, такие как матрица относительных инцидентов положения (PRIM) и матрица частот (FM). После успешного извлечения признаков эти векторы признаков привязываются к нейронной сети для всестороннего обучения и последующей классификации произвольной последовательности белка, полученной в качестве входных данных.
Во многих предыдущих попытках мембранные белки предсказывались через их первичную последовательность аминокислот. Большинство из этих методов было основано на композициях аминокислот (AA) [15] и псевдо-аминокислот (PseAA) [16]. Чоу и Элрод в [15] предложили метод предсказания типа мембранного белка на основе аминокислотного (АА) состава. Эта работа считается пионером в области методов прогнозирования белка на основе аминокислотного состава. Они использовали ковариантно-дискриминантный анализ (CDA) в сочетании с составом AA, который представлял частоты встречаемости AA в первичной последовательности.Проблема с этим методом заключалась в потере информации о последовательности белка, которая напрямую влияла на эффективность модели прогнозирования. Чтобы сохранить порядок последовательности и ее информации, Чжоу предложил композицию PseAA в [16], которая повлияла на улучшение предсказания информации о последовательности белка. Чоу реализовал расширенный CDA, наименьшее расстояние Хэмминга, наименьшее евклидово расстояние [17] и ProtLock [18] с PseAAC, с помощью которого было исследовано улучшение результатов.Cai et al. в [19] использовали композиции PseAA и функционального домена (FD) с машиной опорных векторов (SVM) для предсказания типа мембранного белка. В [20] Cai et al. применены машины опорных векторов с функциями композиции AA.
Эта работа считается пионером в области методов прогнозирования белка на основе аминокислотного состава. Они использовали ковариантно-дискриминантный анализ (CDA) в сочетании с составом AA, который представлял частоты встречаемости AA в первичной последовательности.Проблема с этим методом заключалась в потере информации о последовательности белка, которая напрямую влияла на эффективность модели прогнозирования. Чтобы сохранить порядок последовательности и ее информации, Чжоу предложил композицию PseAA в [16], которая повлияла на улучшение предсказания информации о последовательности белка. Чоу реализовал расширенный CDA, наименьшее расстояние Хэмминга, наименьшее евклидово расстояние [17] и ProtLock [18] с PseAAC, с помощью которого было исследовано улучшение результатов.Cai et al. в [19] использовали композиции PseAA и функционального домена (FD) с машиной опорных векторов (SVM) для предсказания типа мембранного белка. В [20] Cai et al. применены машины опорных векторов с функциями композиции AA.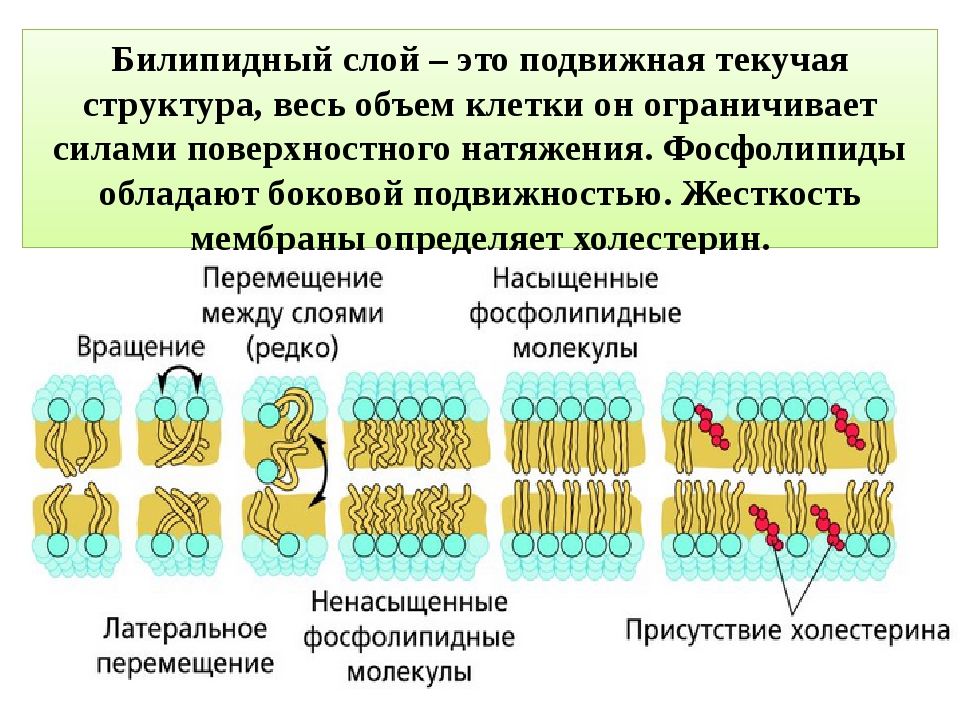 Wang et al. в [21] использовали вариацию SVM с весами для идентификации мембранных белков с использованием особенностей состава PseAA. Контролируемое локально линейное встраивание (SLLE) с классификатором ближайшего соседа было использовано Wang et al. в [22] для выделения признаков и классификации мембранных белков.Чжоу и Цай в [23] использовали паттерны, основанные на амфипатических эффектах порядков последовательности, чтобы облегчить свои существующие методологии. Эти амфифильные эффекты использовались с композицией PseAA, которая ограничивает информацию, относящуюся к гидрофобным и гидрофильным ассоциативным свойствам, и заметно увеличивает процесс прогнозирования типов мембранных белков. Лю и др. в [24, 25] введен анализ спектра Фурье и низкочастотного спектра Фурье на основе композиций PseAA. Основным преимуществом этого анализа было более эффективное использование информации о паттернах белковой последовательности.Chou и Cai в [26] предложили гибридный метод предсказания мембранных белков с использованием GO-PseAA, который был предложен и использован в [27–29], на основе комбинации состава PseAA и генной онтологии (GO).
Wang et al. в [21] использовали вариацию SVM с весами для идентификации мембранных белков с использованием особенностей состава PseAA. Контролируемое локально линейное встраивание (SLLE) с классификатором ближайшего соседа было использовано Wang et al. в [22] для выделения признаков и классификации мембранных белков.Чжоу и Цай в [23] использовали паттерны, основанные на амфипатических эффектах порядков последовательности, чтобы облегчить свои существующие методологии. Эти амфифильные эффекты использовались с композицией PseAA, которая ограничивает информацию, относящуюся к гидрофобным и гидрофильным ассоциативным свойствам, и заметно увеличивает процесс прогнозирования типов мембранных белков. Лю и др. в [24, 25] введен анализ спектра Фурье и низкочастотного спектра Фурье на основе композиций PseAA. Основным преимуществом этого анализа было более эффективное использование информации о паттернах белковой последовательности.Chou и Cai в [26] предложили гибридный метод предсказания мембранных белков с использованием GO-PseAA, который был предложен и использован в [27–29], на основе комбинации состава PseAA и генной онтологии (GO). Этот метод оказался более точным для идентификации пяти типов мембранных белков. Шен и Чоу в [30] представили Оптимизированный теоретико-доказательный классификатор ближайшего соседа (OET-NN) с использованием композиций PseAA, основанных на теории доказательств. Шен и др. в [31] применен алгоритм нечетких ближайших соседей (NN) в сочетании с композициями PseAA.Этот метод был основан на нечеткой математике и дал улучшенный подход в процессе предсказания типа мембранного белка. В [32] Wang et al. предложил новый подход, названный «составным обобщением». В этом методе в качестве мета-классификатора использовались комбинации нескольких классификаторов для повышения эффективности обобщения. Ян и др. в [33] использовали функцию на основе АК и дипептидного состава для своих методов предсказания мембранного белка. Pu et al. в [34] использован Комплексный подход к классификации мембранных белков (IAMPC).Они использовали позиционно-специфичную-оценочную матрицу (PSSM) на основе последовательностей белка AA, которая оказалась лучшим подходом, чем извлечение признаков функционального домена.
Этот метод оказался более точным для идентификации пяти типов мембранных белков. Шен и Чоу в [30] представили Оптимизированный теоретико-доказательный классификатор ближайшего соседа (OET-NN) с использованием композиций PseAA, основанных на теории доказательств. Шен и др. в [31] применен алгоритм нечетких ближайших соседей (NN) в сочетании с композициями PseAA.Этот метод был основан на нечеткой математике и дал улучшенный подход в процессе предсказания типа мембранного белка. В [32] Wang et al. предложил новый подход, названный «составным обобщением». В этом методе в качестве мета-классификатора использовались комбинации нескольких классификаторов для повышения эффективности обобщения. Ян и др. в [33] использовали функцию на основе АК и дипептидного состава для своих методов предсказания мембранного белка. Pu et al. в [34] использован Комплексный подход к классификации мембранных белков (IAMPC).Они использовали позиционно-специфичную-оценочную матрицу (PSSM) на основе последовательностей белка AA, которая оказалась лучшим подходом, чем извлечение признаков функционального домена. Чоу и Шен в [35] реализовали веб-сервер (MemType-2L), который использовался в качестве двухуровневого механизма прогнозирования. Первую фазу использовали для идентификации белковой последовательности как мембранного белка или немембранного белка, а на второй фазе дифференцировали специфический тип мембранного белка. Сервер был основан на функциях, извлеченных с помощью Pse-PSSM (Pseudo-Position-Specific-Scoring-Matrix) с комбинацией классификатора ансамбля.
Чоу и Шен в [35] реализовали веб-сервер (MemType-2L), который использовался в качестве двухуровневого механизма прогнозирования. Первую фазу использовали для идентификации белковой последовательности как мембранного белка или немембранного белка, а на второй фазе дифференцировали специфический тип мембранного белка. Сервер был основан на функциях, извлеченных с помощью Pse-PSSM (Pseudo-Position-Specific-Scoring-Matrix) с комбинацией классификатора ансамбля.
2. Материалы и методы
Набор контрольных данных, используемый в предлагаемой системе для обучения и тестирования, был создан Чжоу и Шен в [35]. Последовательности белков были собраны из версии 51.0, выпущенной 6 октября 2006 г., базы данных Swiss-Prot. Следующие критерии были использованы для сбора высококачественных данных и очень желаемого набора рабочих данных. На первом этапе не включались последовательности с аннотацией типа «фрагмент». Белки с последовательностями менее 50 аминокислотных остатков также не включались.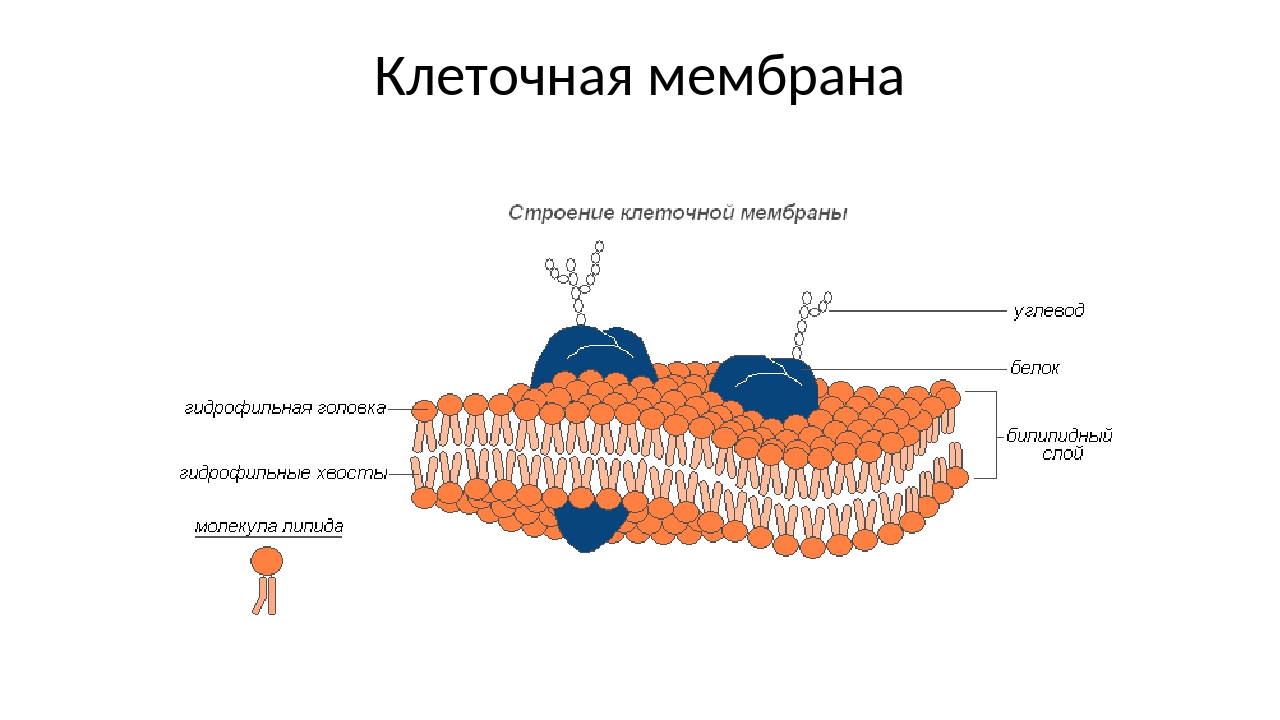 На втором этапе такие последовательности, которые были аннотированы неоднозначными терминами, такими как «потенциальный», «вероятный», «вероятно», «может быть» или «по сходству», не рассматривались как часть этого набора данных. Последовательности, проверенные после описанных выше процедур, сохранялись в наборе данных мембранных белков, если они были помечены термином «мембранный белок», а остальные последовательности, которые не были аннотированы этим термином, сохранялись как набор данных для немембранных белков. Чтобы устранить ошибку гомологии и избыточности, восстановительные последовательности, которые имеют 80% идентичности в последовательности с любыми другими мембранными белками, были исключены.Аналогичная процедура была применена в наборе немембранных данных для удаления избыточных последовательностей. Наконец, был создан набор данных, содержащий 15 547 белковых последовательностей, из которых 7582 были мембранными белками и 7965 были немембранными белками. Этот набор данных является последним эталонным набором данных и в настоящее время используется в основном в прогностических системах, созданных для предсказания мембранных белков.
На втором этапе такие последовательности, которые были аннотированы неоднозначными терминами, такими как «потенциальный», «вероятный», «вероятно», «может быть» или «по сходству», не рассматривались как часть этого набора данных. Последовательности, проверенные после описанных выше процедур, сохранялись в наборе данных мембранных белков, если они были помечены термином «мембранный белок», а остальные последовательности, которые не были аннотированы этим термином, сохранялись как набор данных для немембранных белков. Чтобы устранить ошибку гомологии и избыточности, восстановительные последовательности, которые имеют 80% идентичности в последовательности с любыми другими мембранными белками, были исключены.Аналогичная процедура была применена в наборе немембранных данных для удаления избыточных последовательностей. Наконец, был создан набор данных, содержащий 15 547 белковых последовательностей, из которых 7582 были мембранными белками и 7965 были немембранными белками. Этот набор данных является последним эталонным набором данных и в настоящее время используется в основном в прогностических системах, созданных для предсказания мембранных белков. Чжоу и Шен также подготовили независимый набор тестовых данных из 4333 мембранных белков.
Чжоу и Шен также подготовили независимый набор тестовых данных из 4333 мембранных белков.
2.1. Извлечение признаков
Следующие методы извлечения признаков используются для определения признаков или закономерностей, связанных с любым конкретным белком.Эти методы обсуждаются ниже.
2.1.1. Статистические моменты
Многие предлагаемые методы распознавания образов считают, что статистические моменты полезны для создания признаков из заданного образца, которые не зависят от каких-либо параметров. Многие исследователи использовали моменты, чтобы зафиксировать важные особенности и охарактеризовать функциональные возможности любого заданного паттерна [36, 37]. Моменты – это определенные типы смещенного среднего, которые используются для анализа концентраций некоторых основных конфигураций в задачах, связанных с распознаванием образов.Для различных систем распознавания образов и представлений объектов ортогональные моменты считаются ценным методом. В недавнем исследовании было замечено, что дискретные ортогональные моменты дали лучшие результаты, чем непрерывные ортогональные моменты для дискретных и квантованных данных. Эти дискретные ортогональные моменты обладают способностью преобразовывать представления объектов с минимальной потерей информации [38].
Эти дискретные ортогональные моменты обладают способностью преобразовывать представления объектов с минимальной потерей информации [38].
Чтобы вычислить двумерные моменты, одномерная первичная структура преобразуется в двумерную структуру с использованием схемы основных строк.Размерность двумерной матрицы вычисляется путем извлечения квадратного корня из длины белка, где – размер двумерной квадратной матрицы, а – длина полипептидной цепи.
Существует множество различных форм моментов, которые можно вычислить с помощью любой матрицы или набора векторов, представляющих любой образец. Наиболее распространенными из моментов являются необработанные моменты, которые вычисляются из следующего: исходные моменты принимают исходную точку данных в качестве опорной точки, в то время как компоненты расстояния от начала координат используются для вычисления моментов.Центральные моменты используют центроид данных в качестве опорной точки и вычисляются по следующему уравнению: Здесь и формируют центроид и вычисляются из Одномерные обозначения были преобразованы в обозначения квадратной матрицы, чтобы можно было вычислить моменты Хана.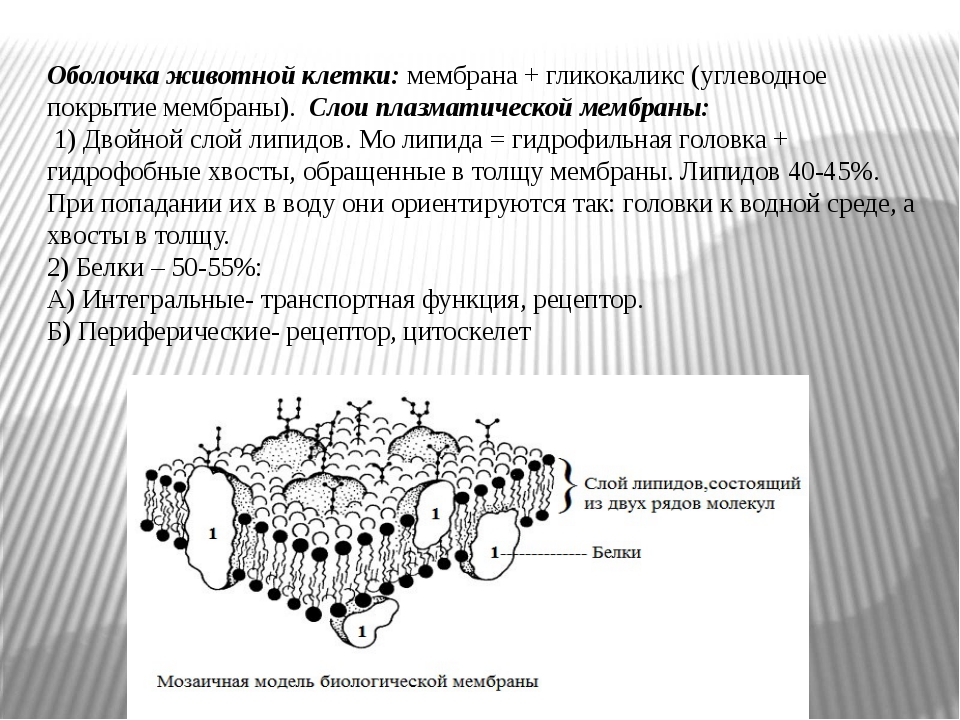 Двумерные моменты Хана – это ортогональные моменты, для которых требуется квадратная матрица в качестве двумерных входных данных. Полином Хана порядка задается как В приведенном выше выражении используется символ Поххаммера, обобщенный как И он упрощается с помощью Гамма-оператора. Необработанные значения моментов Хана обычно масштабируются с использованием весовой функции и квадратной нормы, заданной как Между тем, Ортогональные нормализованные моменты Хана для двух -мерные дискретные данные вычисляются с использованием следующего уравнения: центральные моменты и моменты Хана вычисляются до порядка 3.
Двумерные моменты Хана – это ортогональные моменты, для которых требуется квадратная матрица в качестве двумерных входных данных. Полином Хана порядка задается как В приведенном выше выражении используется символ Поххаммера, обобщенный как И он упрощается с помощью Гамма-оператора. Необработанные значения моментов Хана обычно масштабируются с использованием весовой функции и квадратной нормы, заданной как Между тем, Ортогональные нормализованные моменты Хана для двух -мерные дискретные данные вычисляются с использованием следующего уравнения: центральные моменты и моменты Хана вычисляются до порядка 3.
2.1.2. Позиционно-относительная-инцидентная-матрица (PRIM) и частотная-матрица (FM)
Первым шагом в извлечении признаков является вычисление формирования матрицы входного запроса белка. С этой целью длина белковой последовательности используется для построения PRIM и FM. Эти матрицы затем используются для расчета моментов, посредством которых формируются векторы признаков. Последовательность белка с общими аминокислотными остатками представлена посредством PRIM следующим образом: В данной последовательности белка указание количества остатков в положении th определяется с помощью.В процессе биологической эволюции эта оценка заменяется типом аминокислоты. Значения представляют собой алфавитный порядок 20 природных аминокислот.
Последовательность белка с общими аминокислотными остатками представлена посредством PRIM следующим образом: В данной последовательности белка указание количества остатков в положении th определяется с помощью.В процессе биологической эволюции эта оценка заменяется типом аминокислоты. Значения представляют собой алфавитный порядок 20 природных аминокислот.
После того, как вектор признаков получен, он обучается и классифицируется с помощью многослойной нейронной сети (MLNN) для предсказаний мембранных и немембранных белков.
2.1.3. Нейронные сети с обратным распространением (BP)
В классификации проблем распознавания образов нейронные сети являются одними из наиболее часто используемых методологий.Эти нейросетевые системы являются нелинейно-адаптивными и способны аппроксимировать любую функцию. Алгоритм обучения BP очень хорошо известен для многослойных нейронных сетей с прямой связью и был введен в [39]. Нейронная сеть обратного распространения (BPNN) и нейронная сеть с прямой связью (FFNN) похожи и содержат входной слой, несколько скрытых слоев и выходной слой, как показано на рисунке 1.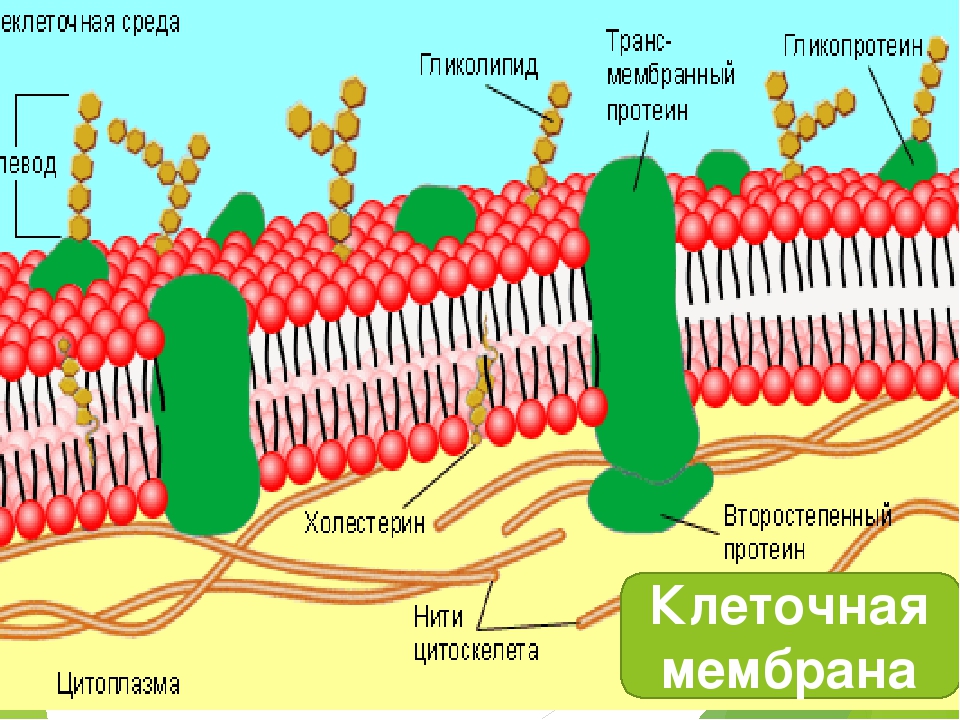 Скрытые слои состоят из выбранного количества нейронов. Эти нейроны действуют как основные обрабатывающие элементы сети.Эти нейроны или узлы образуют созвездие за счет связи между слоями. Через входящие соединения узла он получает взвешенные активации узлов предыдущего уровня. Эти веса суммируются, и результат передается через функцию активации. Результатом этого процесса является активация узла. Затем удельный вес умножается на это значение активации для каждого исходящего соединения и затем передается следующему узлу. Для MLNN используемая функция активации или пороговая функция должна быть нелинейной.Если пороговая функция не является нелинейной, тогда MLNN будет работать как одноуровневая сеть. Наиболее часто применяемый порог или функция активации – это сигмоид логистической функции, который определяется следующей формулой: где – количество заданных входов в сеть. Доступно множество пороговых функций, но это наиболее часто используемая и очень полезная при обучении BPNN. Во время тренировки АД каждый паттерн тренируется по отдельности.
Скрытые слои состоят из выбранного количества нейронов. Эти нейроны действуют как основные обрабатывающие элементы сети.Эти нейроны или узлы образуют созвездие за счет связи между слоями. Через входящие соединения узла он получает взвешенные активации узлов предыдущего уровня. Эти веса суммируются, и результат передается через функцию активации. Результатом этого процесса является активация узла. Затем удельный вес умножается на это значение активации для каждого исходящего соединения и затем передается следующему узлу. Для MLNN используемая функция активации или пороговая функция должна быть нелинейной.Если пороговая функция не является нелинейной, тогда MLNN будет работать как одноуровневая сеть. Наиболее часто применяемый порог или функция активации – это сигмоид логистической функции, который определяется следующей формулой: где – количество заданных входов в сеть. Доступно множество пороговых функций, но это наиболее часто используемая и очень полезная при обучении BPNN. Во время тренировки АД каждый паттерн тренируется по отдельности.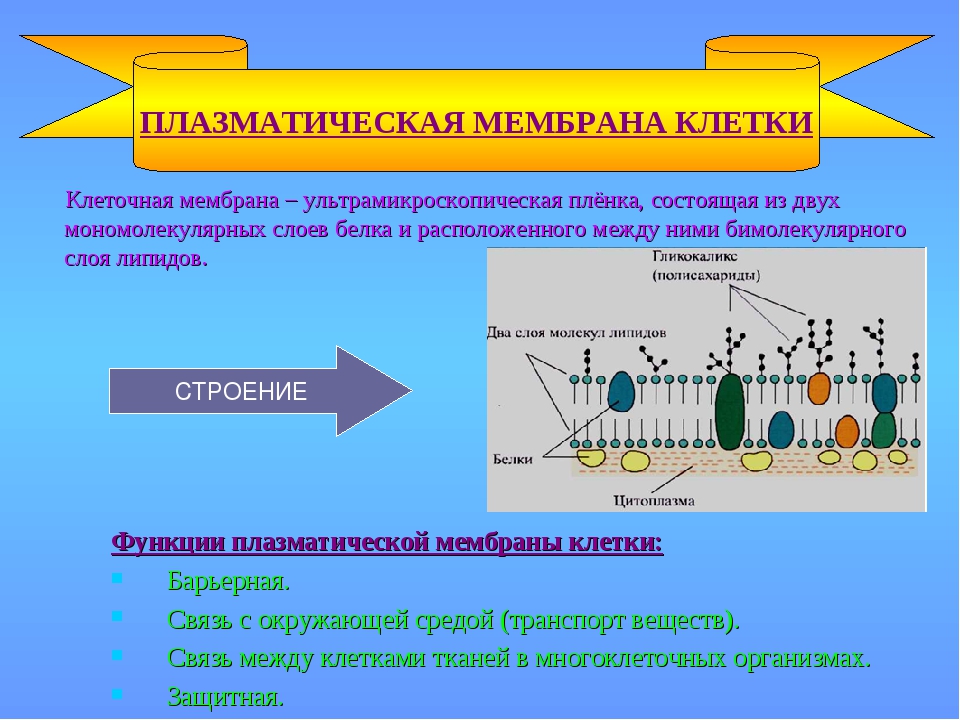 Эпоха – это обучение всех входных шаблонов сети обучающего набора данных.BPNN использует алгоритм достойного градиента. Он использует различные попытки уменьшить свою ошибку по градиенту, чтобы улучшить общую производительность нейронной сети. Эта ошибка может быть выражена как среднеквадратичная ошибка (RMSE), которая формулируется как Здесь сумма ошибок половины средних значений прогнозируемой цели и разности выходных векторов всех шаблонов. В начале тренировки АД веса устанавливаются случайным образом. Веса изменяются в направлении максимального уменьшения и масштабируются с помощью лямбда адаптивной скорости обучения: Свойство производной сигмоидной функции is. Используя вышеупомянутый метод, выполняется вычисление производной логистической сигмоидной функции.Этот метод упрощает вычисления в методе BP. Следовательно, уравнения изменения веса сводятся к следующему: перед применением пороговой функции к необработанной сумме для каждого нейрона значение этой суммы сохраняется. После сохранения этих значений суммы изменения веса выполняются в достаточной степени с использованием основных алгебраических операций.
Эпоха – это обучение всех входных шаблонов сети обучающего набора данных.BPNN использует алгоритм достойного градиента. Он использует различные попытки уменьшить свою ошибку по градиенту, чтобы улучшить общую производительность нейронной сети. Эта ошибка может быть выражена как среднеквадратичная ошибка (RMSE), которая формулируется как Здесь сумма ошибок половины средних значений прогнозируемой цели и разности выходных векторов всех шаблонов. В начале тренировки АД веса устанавливаются случайным образом. Веса изменяются в направлении максимального уменьшения и масштабируются с помощью лямбда адаптивной скорости обучения: Свойство производной сигмоидной функции is. Используя вышеупомянутый метод, выполняется вычисление производной логистической сигмоидной функции.Этот метод упрощает вычисления в методе BP. Следовательно, уравнения изменения веса сводятся к следующему: перед применением пороговой функции к необработанной сумме для каждого нейрона значение этой суммы сохраняется. После сохранения этих значений суммы изменения веса выполняются в достаточной степени с использованием основных алгебраических операций. Алгоритм BP надежен, но не так быстр в обучении. Однако можно изменить различные параметры, чтобы повысить скорость тренировочного процесса в целом.
Алгоритм BP надежен, но не так быстр в обучении. Однако можно изменить различные параметры, чтобы повысить скорость тренировочного процесса в целом.
3. Результаты и обсуждение
Чтобы измерить частоту ошибок в предлагаемой модели классификации для оценки производительности, данные разделены на разделы. Для создания разделов используются самые распространенные статистические методы. Перекрестная проверка – это метод разделения, который обычно используется для проверки эффективности классификации классификатора. В дальнейшем он используется для тестирования с взаимоисключающим разделением складок в наборе данных. Чтобы оценить производительность разработанной модели, необходим некоторый метод, который будет исследовать модель прогнозирования, чтобы проверить, насколько хорошо она работает.Есть несколько параметров, по которым измеряется оценка и оценки эффективности классификаторов. Тип данных и классификация указывают на то, какие параметры следует использовать. Для измерения производительности классификатора обычно используются тесты и инструменты: тест складного ножа, матрица неточностей и кривые ROC.
Для измерения производительности классификатора обычно используются тесты и инструменты: тест складного ножа, матрица неточностей и кривые ROC.
3.1. Тест складного ножа
Валидационный тест складного ножа также известен как перекрестный тест с исключением по одному. В этом тесте данные делятся на -сложные. Экземпляр набора данных тестирования не учитывается, а остальные экземпляры набора данных обучаются с помощью модели классификации.Весь этот процесс выполняется раз. После получения результатов всех прогнозов они дополнительно усредняются для выявления ошибок в оценках. Складной нож – это самый строгий и оперативный тест в перекрестных проверках. Благодаря этому тесту достигаются уникальные результаты. Единственным недостатком при рассмотрении теста складного ножа для оценки является эффективность вычислительных затрат процесса тестирования при выполнении итераций. Этот тест проводился для проверки работоспособности классификатора.Результаты приведены в таблице 1.
| |||||||||||||||||||||||||||||
3.2. Точность
Точность или частота ошибок классификатора используется для измерения его производительности. Степень истинного предсказанного класса классификатора измеряется точностью системы. Это пропорциональность предсказаний, которые верны, по сравнению с ложными. Точности, вычисленные после успешной классификации, также упоминаются в таблице 1, тогда как диаграмма гистограммы для точности показана на рисунке 2.Он сформулирован следующим образом:
3.3. Матрица неточностей
Матрица неточностей в основном рассматривается как инструмент измерения для оценки производительности различных алгоритмов классификации. Результаты классификации сравниваются с фактическими результатами в матрице неточностей. Для представления этой оценки используется матрица, где фактический класс представлен каждым столбцом, а каждая строка – прогнозируемым классом.
В таблице 2 показана таблица непредвиденных обстоятельств или матрица путаницы, которая включает в себя истинные положительные результаты (TP), которые представляют собой истинные результаты, предсказанные как положительные результаты, и истинно отрицательные результаты (TN), которые представляют собой ложные результаты, предсказанные как отрицательные.Ложноположительные результаты (FP) являются ложными, но прогнозируются как положительные, что является ошибкой. Такие типы ошибок известны как ошибки типа I. Кроме того, ложноотрицательные (FN) результаты – это те результаты, которые верны, но прогнозируются как отрицательные. Это также ошибка, известная как ошибка типа II. Матрица путаницы в таблице 3 была получена после успешной классификации мембранных белков от немембранных белков.
| |||||||||||||||||||||||||
3.4. Кривые ROC (приемник-рабочая характеристика)
Для того, чтобы проанализировать основные шестерни методов классификации, был использован тщательный инструмент, а именно кривая ROC, чтобы подчеркнуть точность системы. Точность классификатора можно представить в широком смысле с помощью этого инструмента. Кривая ROC использует график чувствительности (уровень истинных положительных результатов) на вертикальной оси и уровень ложных положительных результатов на горизонтальной оси графика.Работа классификатора считается точной в лучшем случае, когда область кривой находится немного ближе к левому верхнему углу. На рисунке 3 показаны кривые ROC, полученные после выполнения классификаций.
4. Выводы и дальнейшая работа
В этой исследовательской работе предложенные системы дают лучшие результаты для проблемы предсказания мембранного белка. Важность таких систем прогнозирования полезна для определения функциональных возможностей белков, которые имеют важное значение в процессе открытия лекарств.В этой системе использовались различные вычислительные методы, которые ограничены в области распознавания образов. Результаты, полученные в предлагаемой в настоящее время системе, являются более точными и надежными по сравнению с предыдущими результатами из [17, 18] в таблице 1. Поскольку данные о биологической последовательности растут с огромной скоростью в различных базах данных, таких как банк данных Swiss-Prot, остается место для эффективности. и возможности для улучшений в этой области все еще существуют в ближайшем будущем. Содействуя научному сообществу в их экспериментальных целях и студенческому сообществу в их исследовательских целях, мы разработаем удобный веб-сервер, представляющий наш новый метод прогнозирования, представленный в этой статье.
Конфликт интересов
Авторы заявляют об отсутствии конфликта интересов в отношении публикации данной статьи.
Произошла ошибка при настройке пользовательского файла cookie
Этот сайт использует файлы cookie для повышения производительности. Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера для приема файлов cookie
Существует множество причин, по которым cookie не может быть установлен правильно.Ниже приведены наиболее частые причины:
- В вашем браузере отключены файлы cookie. Вам необходимо сбросить настройки своего браузера, чтобы он принимал файлы cookie, или чтобы спросить вас, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались. Чтобы принять файлы cookie с этого сайта, используйте кнопку «Назад» и примите файлы cookie.
- Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
- Дата на вашем компьютере в прошлом.Если часы вашего компьютера показывают дату до 1 января 1970 г., браузер автоматически забудет файл cookie. Чтобы исправить это, установите правильное время и дату на своем компьютере.
- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie. Вы должны отключить приложение при входе в систему или проконсультироваться с системным администратором.
Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу.Чтобы предоставить доступ без файлов cookie потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в файле cookie может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта.Например, сайт не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
Роль мембранных белков в физиологии
Мембранные белки – это белки, которые являются частью клеточных мембран или взаимодействуют с ними, и они несут ответственность за выполнение большинства функций этих мембран.Мембранные белки составляют примерно одну треть белков человека и отвечают за регулирование процессов, которые помогают биологическим клеткам выжить.
Изображение предоставлено: sciencepics / Shutterstock.com
Мембранные белки имеют ряд различных структур и также расположены в разных областях мембраны. Они выполняют широкий спектр функций, и количество белков и типы белков, присутствующих на конкретной мембране, могут варьироваться.
Структура мембранного белка
Клеточные мембраны состоят из двух фосфолипидных бислоев, которые называются листочками.Эти листочки присутствуют на всех клетках, образуя барьер, окружающий каждую клетку. Мембранные белки находятся на этих бислоев фосфолипидов или они взаимодействуют с этими бислоями фосфолипидов.
Есть неполярные мембранные белки, которые являются гидрофобными (водоотталкивающими), и полярные мембранные белки, которые являются гидрофильными (способны смешиваться с водой), которые находятся внутри липидного бислоя. Они напрямую связаны с липидными бислоями, которые создают барьер вокруг каждой клетки.
Интегральные мембранные белки прочно закреплены на мембране.
Белки периферической мембраны не являются постоянной частью мембраны и могут иметь гидрофобные, электростатические и другие нековалентные взаимодействия с мембраной или интегральными белками.
Интегральные белки бывают разных типов, такие как монотопные, битопические, политопные, липидно-заякоренные белки или трансмембранные белки.
Монотопные интегральные белки прикреплены только к одному из двух листочков клетки.
Интегральные белки Bitopic – это трансмембранные белки, которые могут однажды покрывать липидные бислои.Политопные белки также являются трансмембранными белками, которые охватывают липидные бислои более одного раза.
Заякоренный липидом белок имеет ковалентную связь с липидами, которые встроены в фосфолипидный бислой.
Функции мембранного белка
Мембранные белки выполняют широкий спектр функций. К ним относятся:
- Соединения: соединение двух ячеек вместе
Ферментативные функции
Все ферменты представляют собой белок.В результате мембранный белок, внедренный в мембрану, иногда может быть ферментом, активный сайт которого может быть обращен к веществам за пределами липидного бислоя.
Эти типы ферментативных мембранных белков могут работать в группах для выполнения определенных этапов метаболизма, например, расщепления лактозы на углеводы, а затем на моносахариды.
Транспорт
Мембранные белки могут позволить гидрофильным молекулам проходить через клеточную мембрану.Белки транспортной мембраны бывают разных форм, и некоторым требуется энергия для изменения формы и активного перемещения молекул и других веществ через клеточную мембрану. Они делают это, высвобождая АТФ для использования в качестве источника энергии.
- Крепление: становятся точками прикрепления цитоскелета и внеклеточного матрикса
Преобразование сигнала
Некоторые мембранные белки могут иметь сайт связывания. Эти сайты связывания имеют особую форму, которая соответствует форме химического посредника.Например, эти химические посланники могут быть гормонами.
Когда гормон встречается с клеточной стенкой, он соединяется с рецепторным мембранным белком, встроенным в клеточную стенку. Гормон может изменить рецепторный белок и вызвать определенную реакцию, в зависимости от типа гормона или другого вещества, которая будет иметь место внутри клетки.
Распознавание клеток
Другая важная функция мембранных белков – идентификация и распознавание между клетками.Эта конкретная функция полезна для иммунной системы, поскольку она помогает организму распознавать чужеродные клетки, например, которые могут вызывать инфекцию. Гликопротеины – это один из типов мембранных белков, которые могут распознавать клетки.
Межклеточное соединение
Соседние клетки могут иметь мембранные белки, которые соединяются в различных соединениях. Щелевые и плотные соединения.
Эта функция помогает клеткам общаться друг с другом и передавать материалы между собой.
Приложение
Мембранные белки важны для цитоскелета, системы нитей и волокон в цитоплазме клетки и внеклеточного матрикса (ECM), который представляет собой сеть макромолекул, находящихся вне клеток, таких как коллаген, ферменты и гликопротеины. к мембранным белкам.
Присоединение филаментов или волокон в цитоплазме, обнаруженных по всей клетке, может помочь клетке сохранять свою особую форму. Он также сохраняет стабильное расположение мембранных белков.
Присоединение мембранных белков к внеклеточному матриксу может помочь ЕСМ опосредовать изменения, происходящие во внеклеточной и внутриклеточной среде.
Мембранные белки при болезнях
Некоторые заболевания связаны с мутациями мембранных белков. Одним из примеров является мутация под названием V509A, обнаруженная в рецепторе тиреотропина, причем тиреотропин является гормоном, секретируемым гипофизом, который регулирует выработку гормонов щитовидной железы.
Эта мутация увеличивает активность рецептора тиреотропина и приводит к врожденному гипертиреозу, состоянию, которое может вызывать изменения настроения, проблемы со сном и проблемы с желудком.
Другие заболевания, связанные с мутациями в мембранных белках, включают наследственную глухоту, болезнь Шарко-Мари-Тута, которая повреждает периферические нервы за пределами центральной нервной системы, и синдром Дежерина-Соттаса, который влияет на способность человека двигаться.
Изображение предоставлено: Explode / Shutterstock.com
Сводка
Мембранные белкивыполняют ряд важных функций, которые помогают клеткам общаться, поддерживать свою форму, выполнять изменения, вызванные химическими посредниками, а также транспортировать и обмениваться материалами.
Мембранные белки также могут играть роль в прогрессировании заболевания, поскольку иммунная система может использовать мембранные белки для идентификации потенциально вредных чужеродных молекул в организме.
Содержание лекции
Обзор курса
Цели курса
- Опишите основные механизмы и системы, общие для большинства клеток, которые позволяют им выживать, расти и адаптироваться.
- Опишите, как эти механизмы и системы используются и модифицируются в разных ячейках для создания уникальных функций и действий.
- Опишите, как определенные дефекты в этих механизмах и системах приводят к определенным заболеваниям.
- На изображениях под микроскопом определите ключевые структурные особенности клетки и опишите, что их присутствие или отсутствие подразумевает о функции клетки.
- Выявить изменения в клеточной структуре, которые приводят к болезни
Общее понимание того, как клетки работают по отдельности и в группах
Мы опишем основные системы, структуры и пути в клетках, которые позволяют им выживать, расти и адаптироваться.Эти incude
- Общая организация ячеек
- Цитоскелет
- Секреция и эндоцитоз
- Морфология и моторика
- Сотовая связь
- Контроль качества клеток
- Деление клеток и регуляция роста
Сначала мы рассмотрим системы, структуры и пути в общей клетке, а затем исследуем, как они используются и изменяются в определенных типах клеток для создания уникальных функций и действий.Мы выделим клеточные механизмы, разрушение которых приводит к болезни.
В гистологии мы исследуем взаимосвязь между структурой и функцией клеток, чтобы определить, как форма, размер и содержимое клетки способствуют ее функции и активности. Вы узнаете, как определять структурные особенности клеток и как эти подсказки можно использовать для определения функции и типа клетки. Вы также научитесь определять изменения в этих структурных особенностях и лежащих в их основе молекулярных дефектах.Наконец, вы начнете понимать, как эти изменения приводят к болезням.
Зачем изучать клеточную биологию как врач
Клетка – основная единица жизни. Подавляющее большинство живых организмов (например, бактерий и дрожжей) на Земле – это отдельные клетки, которые выживают, растут, размножаются и адаптируются к окружающей среде. Хотя клетки нашего тела более сложные и живут с миллиардами других клеток, они обладают многими из тех же черт, что и одноклеточные организмы. Фактически, некоторые из наших клеток могут при правильных условиях расти и размножаться как отдельные клетки.Например, в 1951 году клетки рака шейки матки были взяты у Генриетты Лакс и выращены как отдельные клетки в культуре. Потомки этих клеток растут сегодня в лабораториях по всему миру. Таким образом, даже в многоклеточных организмах клетка является основной единицей жизни.
Одна из задач биомедицины – понять, как клетки нашего тела вызывают все действия, которые позволяют нам, людям, выживать, расти, размножаться и адаптироваться к окружающей среде. Мы полностью состоим из клеток и материала, который клетки производят, и поэтому все биологические события, необходимые для поддержания нашей жизни, генерируются клетками.Например, когда мы едим, мы всегда подвергаемся риску проглотить токсины. Большинство токсинов, которые мы случайно или намеренно проглатываем, метаболизируются в печени до менее вредных молекул. Если мы исследуем участок печени, чтобы найти, где находится эта детоксицирующая активность, мы обнаружим, что печень содержит в основном большие массы клеток, называемых гепатоцитами. Следовательно, способность к неактивным вредным молекулам должна принадлежать гепатоцитам. Сегодня мы знаем, что каждый из этих гепатоцитов содержит ферменты, которые катализируют химический распад токсинов.
С точки зрения врача, идея о том, что вся деятельность человека опосредуется клетками, означает, что, когда он или она видит болезнь, обычно происходит сбой в каком-то клеточном событии, которое приводит к развитию этой болезни. Например, атеросклероз или утолщение стенок артерий связаны с повышенным уровнем липопротеидов низкой плотности (ЛПНП) в сыворотке крови. ЛПНП извлекается из сыворотки гепатоцитами печени. Мутации, которые снижают способность гепатоцитов поглощать ЛПНП, приводят к более высоким уровням ЛПНП в сыворотке крови, увеличивая риск развития атеросклероза.Знание того, как работают клетки, помогает понять, как возникают болезни. Кроме того, изучая, как клетки поглощают и обрабатывают ЛПНП и регулируют количество холестерина, мы смогли разработать лекарства, которые помогают снизить уровень ЛПНП в сыворотке крови. Таким образом, понимание клеток создает потенциал для разработки методов лечения болезней.
Клеточные мембраны
Функция
Клеточные мембраны действуют как граница, предотвращая потерю критически важного клеточного материала. Сюда входят белки, нуклеиновые кислоты и углеводы, а также строительные блоки для этих макромолекул: аминокислоты, нуклеотиды и сахара соответственно.Кроме того, клеточные мембраны также предотвращают потерю АТФ, клеточной валюты для получения энергии.
Чтобы предотвратить потерю клеточного материала, клеточные мембраны действуют как диффузионный барьер. Все молекулы внутри клетки диффундируют за счет тепловой энергии. Без клеточной мембраны эти молекулы быстро диффундировали бы от клетки и потерялись. Клеточная мембрана образует барьер, через который не могут диффундировать определенные молекулы и химические вещества.
Строение
Структура клеточной мембраны определяет тип молекулы или химических веществ, которые она предотвращает от диффузии.Клеточные мембраны содержат гидрофильные внешние поверхности и гидрофобный внутренний слой. Внешние гидрофильные поверхности позволяют мембранам быть растворимыми в воде, тогда как внутренний гидрофобный слой препятствует прохождению большинства водорастворимых химикатов и молекул.
Мембраны содержат большое количество фосфолипидов. Фосфолипиды имеют две химически различные структуры: гидрофильную головную группу и гидрофобные С-хвосты. В смеси фосфолипидов и воды C-хвосты кластеры, чтобы избежать взаимодействия с водой, тогда как головные группы легко взаимодействуют с водой.Следовательно, наиболее стабильной структурой фосфолипидов в воде является полая сфера. Вода находится снаружи и внутри сферы, а фосфолипиды расположены в двухслойном слое, разделяющем две водные среды. Обратите внимание, что ячейка – это, по сути, большая мембранная сфера с множеством критических компонентов внутри и на поверхности сферы.
Как структура мембраны определяет, каким химическим веществам она препятствует распространению через нее? Плотная упаковка фосфолипидов в мембране предотвращает диффузию более крупных молекул (аминокислот, углеводов).Но даже небольшие ионы, такие как натрий, калий и кальций, не могут диффундировать через клеточную мембрану. В этом случае гидрофобный слой внутри мембраны препятствует диффузии ионов.
Фосфолипиды
Фосфолипиды – это класс молекул, каждый член которого имеет различную структуру и функцию. Фосфолипиды отличаются друг от друга главным образом головными доменами (гидрофильными доменами), тогда как С-хвосты похожи по структуре. Фосфолипиды классифицируются на разные группы в зависимости от структуры и состава головного домена.Подробности обсуждались в биохимии.
Зачем нужны фосфолипиды с разными головными группами? Белки могут различать головные группы. Некоторые белки связываются с головными группами определенных фосфолипидов, позволяя клеткам рекрутировать определенные белки на клеточную мембрану. Важность этого станет более очевидной, когда мы обсудим сотовую связь.
С-хвост фосфолипидов похожи по структуре с одним важным отличием. C-хвосты – это жирные кислоты, содержащие одну или две длинные цепи углеводородов.Отдельные углеводороды могут быть связаны друг с другом одинарной или двойной связью. Цепь со всеми одинарными связями образует прямую углеводородную цепь. Их называют насыщенными липидами. Двойная связь приводит к изгибу цепи, что приводит к изгибу цепи. Их называют ненасыщенными липидами.
Клеточные мембраны содержат смесь насыщенных и ненасыщенных фосфолипидов. Поскольку насыщенные углеводороды являются прямыми, эти фосфолипиды могут упаковываться более плотно. К сожалению, мембрана из всех насыщенных фосфолипидов будет твердой при физиологической температуре.Присутствие ненасыщенных фосфолипидов в мембране создает пространство между фосфолипидами, делая мембрану более жидкой при физиологической температуре.
Не только разные мембраны могут содержать разные типы фосфолипидов, но створки одной мембраны могут отличаться по своему составу фосфолипидов. Например, внешний листок клеточной мембраны обращен к внешней среде и нуждается в определенных типах фосфолипидов. Напротив, внутренний листок клеточной мембраны обращен к цитоплазме клетки и содержит фосфолипиды, которые взаимодействуют с белками.
Мембранные белки
Мембрана, состоящая только из фосфолипидов, могла бы образовывать эффективный диффузионный барьер, но не имела бы многих функций, необходимых для выживания клеток. Например, мембраны ограничивают диффузию большинства небольших молекул, которые необходимы клеткам для выработки энергии и создания более крупных макромолекул. Эти молекулы часто легко доступны вне клетки, но им нужен способ проникнуть через мембрану в клетку. Кроме того, чтобы адаптироваться к изменяющимся условиям, клетки должны ощущать свою внешнюю среду и соответствующим образом реагировать.Мембрана, состоящая только из фосфолипидов, не способна обнаруживать внешние молекулы или условия. Наконец, в многоклеточных организмах клетки должны слипаться друг с другом. Фосфолипиды в мембранах не позволяют образовывать соединения между двумя мембранами.
Для выполнения описанных выше функций мембраны опираются на белки. Белки образуют каналы в мембранах, которые позволяют проходить определенным молекулам или ионам. Белки действуют как рецепторы, обнаруживающие присутствие определенных молекул или ионов во внешней среде.Наконец, белки мембран взаимодействуют с белками других мембран, создавая места прикрепления между мембранами и клетками.
Белки могут связываться с мембранами разными способами. Белки, образующие каналы, рецепторы или точки адгезии, обычно представляют собой интегральные мембранные белки. Эти белки проходят через мембрану хотя бы один раз и могут пересекать мембрану несколько раз (> 10). Эти белки постоянно встроены в мембрану и могут быть удалены только за счет больших затрат энергии или переваривания.
В отличие от интегральных мембранных белков, это белки, ассоциированные с периферией. Эти белки связаны с головными группами определенных фосфолипидов или частями интегральных мембранных белков. Ассоциация этих белков с мембранами носит временный характер. Взаимодействие можно обратить вспять, изменив состав мембраны, морфологию или заряд белка. Эти белки часто обеспечивают структурную поддержку мембран, участвуют в передаче клеточных сигналов или изменяют топологию мембран секреторного пути.
Еще один важный тип взаимодействия между белком и мембраной опосредуется ковалентной связью между белком и фосфолипидом. Взаимодействие происходит между аминокислотой на С-конце белка и головной группой фосфолипида. Эти белки находятся на внешней створке клеточной мембраны, причем белок обращен к внешней среде. Многие из этих белков являются ферментами, которые модифицируют белки в крови или внеклеточном пространстве.
Текучесть мембран
Клеточная мембрана динамична в том смысле, что отдельные фосфолипиды и белки, составляющие мембрану, быстро перемещаются внутри мембраны.Тепловая энергия заставляет липиды и белки диффундировать внутри мембран. Ученые измерили скорость диффузии липидов и белков, и, по одной из оценок, липид может обогнуть бактериальную мембрану за одну секунду.
Диффузионная природа мембран имеет важные биологические и медицинские последствия. Без какой-либо другой системы, действующей на мембрану, липиды и белки в мембране будут случайным образом распределяться по этой мембране. Как мы увидим, клетки затрачивают значительное количество энергии на создание доменов в мембранах, содержащих определенный набор белков, а иногда и липидов.Эти домены играют важную роль в клеточной адгезии и коммуникации.
Структурная опора клеточной мембраны
Клеточным мембранам требуется структурная опора для поддержания формы и предотвращения повреждения фосфолипидного бислоя. В большинстве клеток цитоскелет находится под клеточной мембраной в цитоплазме, обеспечивая структурную поддержку. Актиновые филаменты являются наиболее распространенными, но некоторые клетки используют микротрубочки для образования уникальных структур (например, ресничек). Нити актина образуют сеть нитей под клеточной мембраной.
Цитоскелет обеспечивает структурную поддержку частично за счет взаимодействия с интегральными мембранными белками. Взаимодействие с цитоскелетом ограничивает диффузию мембранных белков и обеспечивает стабильную основу, к которой прикрепляются мембранные белки. Это взаимодействие предотвращает повреждение мембран, когда внешние силы тянут или давят на интегральные мембранные белки.
Клеточная мембрана и потенциал клетки
Совершенно очевидно, почему клеточные мембраны ограничивают диффузию небольших молекул, таких как аминокислоты, АТФ и т. Д.Клетка не хочет терять эти важные молекулы в окружающую среду. Но почему важно, чтобы клеточная мембрана ограничивала диффузию ионов?
Поскольку ионы не могут диффундировать через клеточную мембрану, клетки могут устанавливать различные концентрации ионов в цитозоле по сравнению с внешней средой. Например, внутриклеточная концентрация натрия намного ниже, чем внеклеточная концентрация. Напротив, внутриклеточная концентрация калия намного выше, чем внеклеточная концентрация.Как и в случае натрия, внутриклеточная концентрация кальция очень низкая. Клетки могут устанавливать и поддерживать эти ионные градиенты, потому что ионы не могут диффундировать через мембрану. Ионы могут проходить только через определенные белки, которые образуют каналы или насосы в мембране. Клетки точно регулируют деятельность каналов и насосов. Чистое распределение ионов делает цитозоль клеток электроотрицательным по сравнению с внешней средой.
Какова биологическая роль этих ионных градиентов? Одна из важных ролей – это клеточная коммуникация, в частности, связь между нейронами и их мишенями.Активные нейроны посылают сигналы по своим аксонам, позволяя ионам натрия проникать в цитозоль, тем самым деполяризуя аксон. Клетки также используют ионные градиенты для регулирования активности ферментов. Многим ферментам для своей деятельности необходим кальций. Поскольку цитозольная концентрация кальция низкая, большинство этих ферментов неактивны. Когда клетки хотят активировать эти ферменты, они позволяют цитозольной концентрации кальция повышаться.
Клеточная мембрана – Структура – Фосфолипиды
Клеточные мембраны являются важным компонентом клетки, обеспечивая разделение внутриклеточной и внеклеточной среды.Они состоят из липидов, белков и углеводов.
В этой статье мы рассмотрим основные функции клеточной мембраны, состав мембран и клинические состояния, при которых часть клеточной мембраны является аномальной.
Рис. 1. Структура клеточной мембраны [/ caption]Структура
Упрощенная приблизительная шкала сухого веса приведена в Таблице 1.
| Сухой вес |
| 40% липидов – Э.грамм. молекулы фосфолипидов и холестерин |
| 60% белка – Например. канальные белки и белки-носители |
| 1-10% углеводов – Часто обнаруживается прикрепленным к белкам / липидам на внешней стороне клеточной мембраны – углеводная оболочка, окружающая клетку, часто называется гликокаликсом |
Фосфолипиды
Двухслойная мембрана содержит много видов молекул фосфолипидов с молекулами головы и хвоста разного размера.
Они состоят из головной молекулы, молекулы фосфата, глицерина и двух цепей жирных кислот.
- Головная группа – это полярная группа , например сахар или холин – это означает, что головной конец фосфолипида является гидрофильным.
- Хвост из 2 цепей жирных кислот – обычно состоит из 14-24 атомов углерода (но наиболее распространенные длины атомов углерода – 16 и 18). Если цепь содержит цис-двойную связь, тогда цепь изгибается, что снижает плотную упаковку мембраны и, таким образом, увеличивает ее движение.Поскольку хвост состоит из жирных кислот, он не образует водородных связей с водой и, следовательно, является гидрофобным и неполярным.
Молекулы фосфолипидов, таким образом, амфипатичны, являются как гидрофильными, так и гидрофобными. Они спонтанно образуют в воде бислои, при этом группы головок обращены наружу, а группы хвостов – внутрь.
В бислое существуют силы Ван-дер-Вааль между жирнокислотными хвостами фосфолипида, с электростатическими и водородными связями между гидрофильными группами и водой.
Рис. 2. Диаграмма, показывающая структуру как фосфолипидного бислоя, так и отдельного фосфолипида. [/ caption]Холестерин
Холестерин жизненно важен для многих функций клетки, в том числе, что очень важно, для основной составляющей клеточной мембраны.
Сам холестерин состоит из полярной головки, плоского стероидного кольца и неполярного углеводородного хвоста. Холестерин важен для мембраны, поскольку он помогает поддерживать стабильность и текучесть клеточной мембраны при различных температурах.
Холестерин связывается с соседними молекулами фосфолипидов посредством водородных связей и, следовательно, при низких температурах снижает их упаковку. В целом это означает, что при низких температурах, когда скорость движения самая низкая, сохраняется жидкая фаза .
При высоких температурах холестерин помогает остановить образование кристаллических структур , а жесткое плоское стероидное кольцо предотвращает внутрицепочечную вибрацию и, следовательно, делает мембрану менее текучей.
Мембранные белки
Как показано в таблице выше, типичная клеточная мембрана состоит примерно из 60% белка. Белок так велик, потому что он жизненно важен почти для каждого процесса в клетке. Список всего нескольких функций мембранных белков может включать:
- Катализаторы – ферменты.
- Транспортеры, насосы и ионные каналы.
- Рецепторы гормонов, местных медиаторов и нейромедиаторов.
- Преобразователи энергии.
Более активные клетки или органеллы, например митохондрии, как правило, содержат больше белков, что еще раз показывает, что специализация функций определяет структуру.
Как часть клеточной мембраны, белки могут быть либо глубоко встроены в бислой ( интеграл, ), либо быть ассоциированными с поверхностью клетки ( периферический ).
Функции клеточной мембраны
Клеточные мембраны жизненно важны для нормального функционирования всех клеток нашего тела. В их основные функции входят:
- Образует непрерывный, высокоселективно проницаемый барьер – как вокруг клеток, так и внутриклеточных компартментов.
- Позволяет контролировать закрытую химическую среду – важно для поддержания ионных градиентов.
- Связь – как с внеклеточным, так и с внеклеточным пространством.
- Распознавание – включая распознавание сигнальных молекул, белков адгезии и других клеток-хозяев (очень важно для иммунной системы).
- Генерация сигнала – в ответ на стимул, вызывающий изменение мембранного потенциала.
В клетке разные части мембраны выполняют разные функции, и поэтому их структура предназначена для этого.Пример этой специализации можно увидеть в разных частях нерва; клеточная мембрана в аксоне специализирована для электропроводности, тогда как конец нерва специализирован для синапсов, то есть состав мембраны отличается.
[старт-клиника]
Клиническая значимость – Наследственный сфероцитоз
Наследственный сфероцитоз – это состояние, при котором спектрин, – белок периферического цитоскелета, истощены на 40-80%.Существуют как аутосомно-доминантные, так и рецессивные формы заболевания разной степени тяжести. В результате отсутствия спектрина эритроциты не могут эффективно поддерживать свою двояковогнутую структуру и принимают сферическую форму. Это снижает их способность проходить через микрососуды тела и приводит к усилению лизиса эритроцитов. Есть 3 других типа сфероцитоза, которые возникают в результате дефектов анкирина, полосы 3 и белка 4.2, однако спектрин является наиболее значимым.
Признаки и симптомы этого состояния включают:
- От легкой до средней анемия
- Возможна желтуха
- Возможна спленомегалия Рис. 3. Схема мазка периферической крови пациента с наследственным сфероцитозом. [/ caption]
[конец клинической]
|