Клеточная мембрана | Биология
Клеточная мембрана также называется плазматической (или цитоплазматической) мембраной и плазмалеммой. Данная структура не только отделяет внутреннее содержимое клетки от внешней среды, но также входит с состав большинства клеточных органелл и ядра, в свою очередь отделяя их от гиалоплазмы (цитозоля) — вязко-жидкой части цитоплазмы. Договоримся называть цитоплазматической мембраной ту, которая отделяет содержимое клетки от внешней среды. Остальными терминами обозначать все мембраны.
Строение клеточной мембраны
В основе строения клеточной (биологической) мембраны лежит двойной слой липидов (жиров). Формирование такого слоя связано с особенностями их молекул. Липиды не растворяются в воде, а по-своему в ней конденсируются. Одна часть отдельно взятой молекулы липида представляет собой полярную головку (она притягивается водой, т. е. гидрофильна), а другая — пару длинных неполярных хвостов (эта часть молекулы отталкивается от воды, т. е. гидрофобна). Такое строение молекул заставляет их «прятать» хвосты от воды и поворачивать к воде свои полярные головки.
В результате образуется двойной липидный слой, в котором неполярные хвосты находятся внутри (обращены друг к другу), а полярные головки обращены наружу (к внешней среде и цитоплазме). Поверхность такой мембраны гидрофильна, а внутри она гидрофобна.

В клеточных мембранах среди липидов преобладают фосфолипиды (относятся к сложным липидам). Их головки содержат остаток фосфорной кислоты. Кроме фосфолипидов есть гликолипиды (липиды + углеводы) и холестерол (относится к стеролам). Последний придает мембране жесткость, размещаясь в ее толще между хвостами остальных липидов (холестерол полностью гидрофобный).
За счет электростатического взаимодействия, к заряженным головкам липидов присоединяются некоторые молекулы белков, которые становятся поверхностными мембранными белками. Другие белки взаимодействуют с неполярными хвостами, частично погружаются в двойной слой или пронизывают его насквозь.
Таким образом, клеточная мембрана состоит из двойного слоя липидов, поверхностных (периферических), погруженных (полуинтегральных) и пронизывающих (интегральных) белков. Кроме того, некоторые белки и липиды с внешней стороны мембраны связаны с углеводными цепями.

Это жидкостно-мозаичная модель строения мембраны была выдвинута в 70-х годах XX века. До этого предполагалась бутербродная модель строения, согласно которой липидный бислой находится внутри, а с внутренней и наружной стороны мембрана покрыта сплошными слоями поверхностных белков. Однако накопление экспериментальных данных опровергло эту гипотезу.
Толщина мембран у разных клеток составляет около 8 нм. Мембраны (даже разные стороны одной) отличаются между собой по процентному соотношению различных видов липидов, белков, ферментативной активности и др. Какие-то мембраны более жидкие и более проницаемые, другие более плотные.
Разрывы клеточной мембраны легко сливаются из-за физико-химических особенностей липидного бислоя. В плоскости мембраны липиды и белки (если только они не закреплены цитоскелетом) перемещаются.
Функции клеточной мембраны
Большинство погруженных в клеточную мембрану белков выполняют ферментативную функцию (являются ферментами). Часто (особенно в мембранах органоидов клетки) ферменты располагаются в определенной последовательности так, что продукты реакции, катализируемые одним ферментом, переходят ко второму, затем третьему и т. д. Образуется конвейер, который стабилизируют поверхностные белки, т. к. не дают ферментам плавать вдоль липидного бислоя.
Клеточная мембрана выполняет отграничивающую (барьерную) от окружающей среды и в то же время транспортную функции.
Можно сказать, это ее самое главное назначение. Цитоплазматическая мембрана, обладая прочностью и избирательной проницаемостью, поддерживает постоянство внутреннего состава клетки (ее гомеостаз и целостность).При этом транспорт веществ происходит различными способами. Транспорт по градиенту концентрации предполагает передвижение веществ из области с их большей концентрацией в область с меньшей (диффузия). Так, например, диффундируют газы (CO2, O2).
Бывает также транспорт против градиента концентрации, но с затратой энергии.
Транспорт бывает пассивным и облегченным (когда ему помогает какой-нибудь переносчик). Пассивная диффузия через клеточную мембрану возможна для жирорастворимых веществ.
Есть особые белки, делающие мембраны проницаемыми для сахаров и других водорастворимых веществ. Такие переносчики соединяются с транспортируемыми молекулами и протаскивают их через мембрану. Так переносится глюкоза внутрь эритроцитов.
Пронизывающие белки, объединяясь, могут образовывать пору для перемещения некоторых веществ через мембрану. Такие переносчики не перемещаются, а образуют в мембране канал и работают аналогично ферментам, связывая определенное вещество. Перенос осуществляется благодаря изменению конформации белка, благодаря чему в мембране образуются каналы. Пример — натрий-калиевый насос.
Транспортная функция клеточной мембраны эукариот также реализуется за счет эндоцитоза (и экзоцитоза). Благодаря этим механизмам в клетку (и из нее) попадают крупные молекулы биополимеров, даже целые клетки. Эндо- и экзоцитоз характерны не для всех клеток эукариот (у прокариот его вообще нет). Так эндоцитоз наблюдается у простейших и низших беспозвоночны; у млекопитающих лейкоциты и макрофаги поглощают вредные вещества и бактерии, т. е. эндоцитоз выполняет защитную функцию для организма.
Эндоцитоз делится на фагоцитоз (цитоплазма обволакивает крупные частицы) и пиноцитоз (захват капелек жидкости с растворенными в ней веществами). Механизм этих процессов приблизительно одинаков. Поглощаемые вещества на поверхности клеток окружаются мембраной. Образуется пузырек (фагоцитарный или пиноцитарный), который затем перемещается внутрь клетки.
Экзоцитоз — это выведение цитоплазматической мембраной веществ из клетки (гормонов, полисахаридов, белков, жиров и др.). Данные вещества заключаются в мембранные пузырьки, которые подходят к клеточной мембране. Обе мембраны сливаются и содержимое оказывается за пределами клетки.
Цитоплазматическая мембрана выполняет рецепторную функцию. Для этого на ее внешней стороне располагаются структуры, способные распознавать химический или физический раздражитель. Часть пронизывающих плазмалемму белков с наружней стороны соединены с полисахаридными цепочками (образуя гликопротеиды). Это своеобразные молекулярные рецепторы, улавливающие гормоны. Когда конкретный гормон связывается со своим рецептором, то изменяет его структуру. Это в свою очередь запускает механизм клеточного ответа. При этом могут открываться каналы, и в клетку могут начать поступать определенные вещества или выводиться из нее.
Рецепторная функция клеточных мембран хорошо изучена на основе действия гормона инсулина. При связывании инсулина с его рецептором-гликопротеидом происходит активация каталитической внутриклеточной части этого белка (фермента аденилатциклазы). Фермент синтезирует из АТФ циклическую АМФ. Уже она активирует или подавляет различные ферменты клеточного метаболизма.
Рецепторная функция цитоплазматической мембраны также включает распознавание соседних однотипных клеток. Такие клетки прикрепляются друг к другу различными межклеточными контактами.
В тканях с помощью межклеточных контактов клетки могут обмениваться между собой информацией с помощью специально синтезируемых низкомолекулярных веществ. Одним из примеров подобного взаимодействия является контактное торможение, когда клетки прекращают рост, получив информацию, что свободное пространство занято.
Межклеточные контакты бывают простыми (мембраны разных клеток прилегают друг к другу), замковыми (впячивания мембраны одной клетки в другую), десмосомы (когда мембраны соединены пучками поперечных волокон, проникающих в цитоплазму). Кроме того, есть вариант межклеточных контактов за счет медиаторов (посредников) — синапсы. В них сигнал передается не только химическим, но и электрическим способом. Синапсами передаются сигналы между нервными клетками, а также от нервных к мышечным.
biology.su
Клеточная мембрана – строение и функции органоида
Снаружи клетка покрыта плазматической мембраной (или наружной клеточной мембраной) толщиной около 6-10нм.
Строение клеточной мембраны
Клеточная мембрана это плотные пленки из белков и липидов (в основном, фосфолипидов). Молекулы липидов расположены упорядоченно — перпендикулярно к поверхности, в два слоя, так, что их части, интенсивно взаимодействующие с водой (гидрофильные), направлены наружу, а части, инертные к воде (гидрофобные) — внутрь.
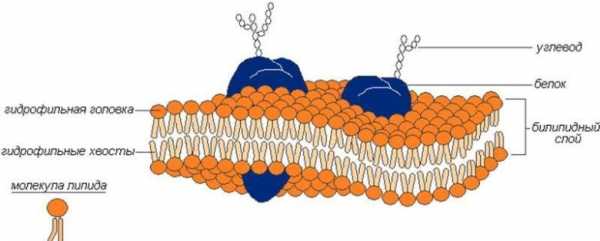 Строение клеточной мембраны
Строение клеточной мембраныМолекулы белка расположены несплошным слоем на поверхности липидного каркаса с обеих его сторон. Часть их погружена в липидный слой, а некоторые проходят через него насквозь, образуя участки, проницаемые для воды. Эти белки выполняют различные функции — одни из них являются ферментами, другие — транспортными белками, участвующими в переносе некоторых веществ из окружающей среды в цитоплазму и в обратном направлении.
Основные функции клеточной мембраны
Одним из основных свойств биологических мембран является избирательная проницаемость (полупроницаемость) — одни вещества проходят через них с трудом, другие легко и даже в сторону большей концентрации Так, для большинства клеток концентрация ионов Na внутри значительно ниже, чем в окружающей среде. Для ионов K характерно обратное соотношение: их концентрация внутри клетки выше, чем снаружи. Поэтому ионы Na всегда стремятся проникнуть в клетку, а ионы K — выйти наружу. Выравниванию концентраций этих ионов препятствует присутствие в мембране особой системы, играющей роль насоса, который откачивает ионы Na из клетки и одновременно накачивает ионы K внутрь.
Стремление ионов Na к перемещению снаружи внутрь используется для транспорта сахаров и аминокислот внутрь клетки. При активном удалении ионов Na из клетки создаются условия для поступления глюкозы и аминокислот внутрь ее.
 Транспорт через клеточную мембрану
Транспорт через клеточную мембрануУ многих клеток поглощение веществ происходит также путем фагоцитоза и пиноцитоза. При фагоцитозе гибкая наружная мембрана образует небольшое углубление, куда попадает захватываемая частица. Это углубление увеличивается, и, окруженная участком наружной мембраны, частица погружается в цитоплазму клетки. Явление фагоцитоза свойственно амебам и некоторым другим простейшим, а также лейкоцитам (фагоцитам). Аналогично происходит и поглощение клетками жидкостей, содержащих необходимые клетке вещества. Это явление было названо
Наружные мембраны различных клеток существенно отличаются как по химическому составу своих белков и липидов, так и по их относительному содержанию. Именно эти особенности определяют разнообразие в физиологической активности мембран различных клеток и их роль, в жизнедеятельности клеток и тканей.
С наружной мембраной связана эндоплазматическая сеть клетки. При помощи наружных мембран осуществляются различные типы межклеточных контактов, т.е. связь между отдельными клетками.
Для многих типов клеток характерно наличие на их поверхности большого количества выступов, складок, микроворсинок. Они способствуют как значительному увеличению площади поверхности клеток и улучшению обмена веществ, так и более прочным связям отдельных клеток друг с другом.
У растительных клеток снаружи клеточной мембраны имеются толстые, хорошо различимые в оптический микроскоп оболочки, состоящие из клетчатки (целлюлозы). Они создают прочную опору растительным тканям (древесина).
Некоторые клетки животного происхождения тоже имеют ряд внешних структур, находящихся поверх клеточной мембраны и имеющих защитный характер. Примером может быть хитин покровных клеток насекомых.
Функции клеточной мембраны (кратко)
| Функция | Описание |
|---|---|
| Защитный барьер | Отделяет внутренние органеллы клетки от внешней среды |
| Регулирующая | Производит регуляцию обмена веществ между внутренним содержимым клетки и наружной средой |
| Разграничивающая (компартментализация) | Разделение внутреннего пространства клетки на независимые блоки (компартменты) |
| Энергетическая | – Накопление и трансформация энергии; – световые реакции фотосинтеза в хлоропластах; – Всасывание и секреция. |
| Рецепторная (информационная) | Участвует в формировании возбуждения и его проведения. |
| Двигательная | Осуществляет движение клетки или отдельных ее частей. |
animals-world.ru
Какие функции выполняет наружная клеточная мембрана? Строение наружной клеточной мембраны
Изучением строения клеток прокариотических организмов, а также растений животных и человека занимается раздел биологии, называемый цитологией. Ученые установили, что содержимое клетки, которое находится внутри нее, построено довольно сложно. Его окружает так называемый поверхностный аппарат, в состав которого входят наружная клеточная мембрана, надмембранные структуры: гликокаликс и клеточная стенка, а также микронити, пеликула и микротрубочки, образующие её подмембранный комплекс.
В данной статье мы изучим строение и функции наружной клеточной мембраны, входящей в поверхностный аппарат различных видов клеток.

Какие функции выполняет наружная клеточная мембрана
Как было описано ранее, наружная мембрана является частью поверхностного аппарата каждой клетки, который успешно отделяет ее внутреннее содержимое и защищает клеточные органеллы от неблагоприятных условий внешней среды. Еще одна функция – это обеспечение обмена веществ между клеточным содержимым и тканевой жидкостью, поэтому наружная клеточная мембрана осуществляет транспорт молекул и ионов, поступающих в цитоплазму, а также помогает удалять шлаки и избыток токсичных веществ из клетки.

Строение клеточной мембраны
Мембраны, или плазмалеммы различных типов клеток сильно отличаются между собой. Главным образом, химическим строением, а также относительным содержанием в них липидов, гликопротеидов, белков и, соответственно, характером рецепторов, находящихся в них. Наружная клеточная мембрана, строение и функции которой определяются прежде всего индивидуальным составом гликопротеидов, берет участие в распознавании раздражителей внешней среды и в реакциях самой клетки на их действия. С белками и гликолипидами клеточных мембран могут взаимодействовать некоторые виды вирусов, вследствие чего они проникают в клетку. Вирусы герпеса и гриппа могут использовать плазмалемму клетки-хозяина для построения свой защитной оболочки.

А вирусы и бактерии, так называемые бактериофаги, прикрепляются к мембране клетки и в месте контакта растворяют ее с помощью особого фермента. Затем в образовавшееся отверстие проходит молекула вирусной ДНК.
Особенности строения плазмалеммы эукариот
Напомним, что наружная клеточная мембрана выполняет функцию транспорта, то есть переноса веществ в цитоплазму клетки и из нее во внешнюю среду. Для осуществления такого процесса необходимо специальное строение. Действительно, плазмалемма представляет собой постоянную, универсальную для всех эукариотических клеток систему поверхностного аппарата. Это тоненькая (2-10 Нм), но достаточно плотная многослойная пленка, которая покрывает всю клетку. Её строение было изучено в 1972 году такими учеными, как Д. Сингер и Г. Николсон, ими же создана жидкостно-мозаичная модель клеточной мембраны.
Главные химические соединения, которые её образуют – это упорядоченно расположенные молекулы белков и определенных фосфолипидов, которые вкраплены в жидковатую липидную среду и напоминают мозаику. Таким образом, клеточная мембрана состоит из двух слоев липидов, неполярные гидрофобные «хвосты» которых находятся внутри мембраны, а полярные гидрофильные головки обращены к цитоплазме клетки и к межклеточной жидкости.
Слой липидов пронизывается крупными белковыми молекулами, образующими гидрофильные поры. Именно через них транспортируются водные растворы глюкозы и минеральных солей. Некоторые белковые молекулы находятся как на внешней, так и на внутренней поверхности плазмалеммы. Таким образом, на наружной клеточной мембране в клетках всех организмов, имеющих ядра, находятся молекулы углеводов, связанные ковалентными связями с гликолипидами и гликопротеидами. Содержание углеводов в клеточных мембранах колеблется от 2 до 10%.

Строение плазмалеммы прокариотических организмов
Наружная клеточная мембрана у прокариот выполняет сходные функции с плазмалеммами клеток ядерных организмов, а именно: восприятие и передача информации, поступающей из внешней среды, транспорт ионов и растворов в клетку и из нее, защита цитоплазмы от чужеродных реагентов извне. Она может образовывать мезосомы – структуры, возникающие при впячивании плазмалеммы внутрь клетки. На них могут находиться ферменты, участвующие в метаболических реакциях прокариот, например, в репликации ДНК, синтезе белков.
Мезосомы также содержат окислительно-восстановительные ферменты, а у фотосинтетиков находятся бактериохлорофилл (у бактерий) и фикобилин (у цианобактерий).
Роль наружных мембран в межклеточных контактах
Продолжая отвечать на вопрос, какие функции выполняет наружная клеточная мембрана, остановимся на ее роли в межклеточных контактах. У растительных клеток в стенках наружной клеточной мембраны образуются поры, переходящие в целлюлозный слой. Через них возможен выход цитоплазмы клетки наружу, такие тонкие каналы называют плазмодесмами.

Благодаря им связь между соседними растительными клетками очень прочная. У клеток человека и животных места контактов соседних клеточных мембран называются десмосомами. Они характерны для эндотелиальных и эпителиальных клеток, а также встречаются у кардиомиоцитов.
Вспомогательные образования плазмалеммы
Разобраться, чем отличаются растительные клетки от животных, помогает изучение особенностей строения их плазмалемм, которые зависят от того, какие функции выполняет наружная клеточная мембрана. Над ней у животных клеток находится слой гликокаликс. Он образован молекулами полисахаридов, связанных с белками и липидами наружной клеточной мембраны. Благодаря гликокаликсу между клетками возникает адгезия (слипание), приводящая к образованию тканей, поэтому он принимает участие в сигнальной функции плазмалеммы – распознавании раздражителей внешней среды.
Как осуществляется пассивный транспорт определенных веществ через клеточные мембраны
Как было уже сказано ранее, наружная клеточная мембрана участвует в процессе транспортировки веществ между клеткой и внешней средой. Существует два вида переноса через плазмалемму: пассивный (дифузионный) и активный транспорт. К первому относится диффузия, облегченная диффузия и осмос. Движение веществ по градиенту концентрации зависит, прежде всего, от массы и величины молекул, проходящих через клеточную мембрану. Например, мелкие неполярные молекулы легко растворяются в среднем липидном слое плазмалеммы, продвигаются через нее и оказываются в цитоплазме.

Крупные молекулы органических веществ проникают в цитоплазму с помощью специальных белков-переносчиков. Они имеют видовую специфичность и, соединяясь с частицей или ионом, без затрат энергии пассивно переносят их через мембрану по градиенту концентрации (пассивный транспорт). Этот процесс лежит в основе такого свойства плазмалеммы, как избирательная проницаемость. В процессе пассивного транспорта энергия молекул АТФ не используется, и клетка сберегает её на другие метаболические реакции.
Активный транспорт химических соединений через плазмалемму
Так как наружная клеточная мембрана обеспечивает перенос молекул и ионов из внешней среды внутрь клетки и обратно, становится возможным вывод продуктов диссимиляции, являющихся токсинами, наружу, то есть в межклеточную жидкость. Активный транспорт происходит против градиента концентрации и требует использования энергии в виде молекул АТФ. В нем также участвуют белки-переносчики, называемые АТФ-азами, являющиеся одновременно и ферментами.

Примером такого транспорта служит натрий-калиевый насос (ионы натрия переходят из цитоплазмы во внешнюю среду, а ионы калия закачиваются в цитоплазму). К нему способны эпителиальные клетки кишечника и почек. Разновидностями такого способа переноса служат процессы пиноцитоза и фагоцитоза. Таким образом, изучив, какие функции выполняет наружная клеточная мембрана, можно установить, что к процессам пино- и фагоцитоза способны гетеротрофные протисты, а также клетки высших животных организмов, например, лейкоциты.
Биоэлектрические процессы в клеточных мембранах
Установлено, что существует разность потенциалов между наружной поверхностью плазмалеммы (она заряжена положительно) и пристеночным слоем цитоплазмы, заряженным отрицательно. Ее назвали потенциалом покоя, и она присуща всем живым клеткам. А нервная ткань имеет не только потенциал покоя, но и способна к проведению слабых биотоков, которое называют процессом возбуждения. Наружные мембраны нервных клеток-нейронов, принимая раздражение от рецепторов, начинают менять заряды: ионы натрия массированно поступают внутрь клетки и поверхность плазмалеммы становится электроотрицательной. А пристеночный слой цитоплазмы вследствие избытка катионов получает положительный заряд. Это объясняет, по какой причине происходит перезарядка наружной клеточной мембраны нейрона, что вызывает проведение нервных импульсов, лежащих в основе процесса возбуждения.
fb.ru
Мембрана наружная – это… Что такое Мембрана наружная?
- Мембрана наружная
- Мембрана наружная
внешний слой клеточной стенки (см.) грам- бактерий. Основой М. н. являются липополисахаридный и липопротеидный слои, формирующие матрицу, в которой заключены специфические (матричные) белки. Молекулы 2 матричных белков (поринов) в соединении с липопротеином проникают через оба слоя, соединяются нековалентно с пептидогликаном и образуют канальцы с проходящими по ним в цитоплазму небольшими гидрофильными молекулами. Наружная поверхность М. н мозаична: в поля липопротеида вкраплены молекулы белков и липополисахарида. М. н. выполняет функции каркаса, барьера, специфического транспорта и простой диффузии веществ; протеины служат рецепторами для фагов, участвуют в конъюгации и контролируют деление клетки
(Источник: «Словарь терминов микробиологии»)
.
- Мелиоидоз
- Мембрана ундулирующая
Смотреть что такое “Мембрана наружная” в других словарях:
Наружная запирательная мышца — Наружная запирательная мышца … Википедия
Мембрана клеточная — (лат. мембрана кожица) биологическая «кожица», окружающая протоплазму живой клетки (см. Клетка). Участвует в регуляции обмена веществ между клеткой и окружающей её средой. У некоторых клеток клеточная мембрана единственная структура, служащая… … Концепции современного естествознания. Словарь основных терминов
Ядерная оболочка я мембрана — Ядерная оболочка, я. мембрана * ядзерная абалонка, я. мембрана * nuclear envelope or n. membrane or karyotheca or karyolemma двойная липопротеидная мембрана, которая окружает ядра эукариотических клеток, отделяя их от цитоплазмы. Внешняя… … Генетика. Энциклопедический словарь
пограничная мембрана глиальная наружная — (m. l. glialis externa, LNH) П. м., образованная нейроглией, отделяющая слой палочек и колбочек сетчатки от наружного зернистого слоя … Большой медицинский словарь
Митохондрия — Электронномикроскопическая фотография, показывающая митохондрии млекопитающего в поперечном сечении Митохондрия (от … Википедия
Клеточная стенка (оболочка) бактерий — структура бактерий и грибов, располагающаяся между цитоплазматической мембраной и капсулой (если таковая имеется) или ионизированным слоем внешней среды. Защищает бактерии от осмотического шока (10 25 атм и более) и др. факторов, определяет форму … Словарь микробиологии
плазмалемма — наружная цитоплазматическая мембрана, отделяющая цитоплазму от клеточной стенки. Участвует в обмене веществ между цитоплазмой и внешней средой и в построении клеточной стенки … Анатомия и морфология растений
Куртка штормовая — (штормовка) верхний слой одежды туристов и альпинистов. Она призвана защищать от ветра и влаги. При этом желательно чтобы испарения от тела человека выводились наружу. Штормовка должна быть максимально лёгкой и компактной. Содержание 1… … Энциклопедия туриста
Кровено́сные сосу́ды — (vasa sanguifera, vaea sanguinea) образуют замкнутую систему, по которой осуществляется транспорт крови от сердца на периферию ко всем органам и тканям и обратно к сердцу. Артерии несут кровь от сердца, а по венам кровь возвращается к сердцу.… … Медицинская энциклопедия
Поверхностный слой — Длинный лучевой разгибатель запястья (m. extensor carpi radialis longus) (рис. 90, 113, 114, 116, 118, 122, 123, 125) сгибает пред плечье в локтевом суставе, разгибает кисть и принимает участие в ее отведении. Мышца имеет веретенообразную форму и … Атлас анатомии человека
dic.academic.ru
Внешняя бактериальная мембрана — Википедия Переиздание // WIKI 2

Подробная схема строения клеточной стенки грамотрицательных бактерий
Вне́шняя бактериа́льная мембра́на, или нару́жная бактериа́льная мембра́на (англ. bacterial outer membrane) — биологическая мембрана, располагающаяся поверх слоя пептидогликана у грамотрицательных бактерий. По составу она отличается от внутренней, клеточной мембраны. На её поверхности находятся липополисахариды, являющиеся антигенами грамотрицательных патогенных бактерий.
Состав

Строение липополисахарида
Внешняя мембрана, как и клеточная, имеет толщину 7—8 нм[1]. Подобно клеточной мембране, внешняя мембрана представляет собой липидный бислой, который состоит из фосфолипидов, преимущественно фосфатидилэтаноламина, фосфатидилглицерола[en] и дифосфатидилглицерола. Однако во внешней мембране фосфолипиды распределены между слоями несимметрично: внешний лист мембраны образован преимущественно липополисахаридами. Исключениями являются цианобактерии и виды рода Neisseria, у которых во внешнем слое мембраны одновременно присутствуют фосфолипиды и липоолигосахариды. В структуре липополисахарида можно выделить четыре структурных блока:
Внешняя мембрана богата белками, их обозначают Omp[en] (от англ. outer membrane protein). Белки могут покрывать до половины поверхности внешней мембраны. Белки Omp подразделяют на главные и минорные. Главные Omp-белки синтезируются постоянно; к их числу относятся липопротеин Брауна, липопротеины спирохет, OmpA и неспецифический порин[en] OmpF[3].
В структурно-функциональном отношении все белки внешней мембраны можно подразделить на три группы:
Иногда на препаратах обнаруживаются зоны контакта клеточной и внешней мембран. Такие участки называют контактами Байера. Вероятно, непосредственного контакта между мембранами не происходят, и они связываются друг с другом специальными белками в образующейся на месте контакте бреши в пептидогликановом слое[4].
Функции
Во внешней мембране имеются постоянно открытые ионные каналы, из-за чего на ней не может поддерживаться постоянный электрохимический градиент ионов натрия или протонов, поэтому внешняя мембрана не принимает участия в метаболизме клетки. Она также не имеет отношения к биосинтезу белков, липидов и полисахаридов, хотя может быть задействована в их секреции. Однако во внешней мембране находятся некоторые ферменты — пермеазы, гидролазы, иногда — окислительные ферменты[en], такие как марганец-оксидаза[1].
Функции внешней мембраны в бактериальной клетке очень разнообразны. Она совместно с клеточной мембраной образует периплазматическое пространство (периплазму), наряду с клеточной стенкой придаёт жёсткость клетке, служит фильтром, который не даёт попасть в периплазму крупным гидрофильным молекулам и задерживает гидрофобные молекулы. Она не даёт покинуть клетку многим ферментам периплазмы, участвует в поступлении в клетку питательных веществ, а также выделении наружу антибиотиков, токсинов, метаболитов и разнообразных белков. Внешняя мембрана опосредует неспецифичную адгезию бактериальных клеток, взаимодействует с бактериофагами, поверхностными рецепторами как прокариотических, так и эукариотических клеток, а также антителами. Благодаря внешней мембране жёлчные кислоты и другие амфифильные детергенты, а также антибиотики действуют на грамотрицательные бактерии слабее, чем на грамположительные[5]. Липоолигосахариды — важнейшие антигены патогенных грамотрицательных бактерий, их также называют эндотоксинами[3].
Биогенез
Механизмы, которые обеспечивают доставку компонентов внешней мембраны на поверхность клетки, до конца не ясны. Компоненты липополисахарида — липид A и O-антигеновые повторяющиеся единицы — синтезируются на цитоплазматической стороне клеточной мембраны и доставляются наружу независимо двумя специализированными транспортными системами, а именно, переносчиком O-антигена Wzx (RfbX) и ABC-транспортера[en] MsbA, который перемещает липид A с внутреннего липидного слоя клеточной мембраны в наружный[6][7][8][9][10]. Полимеризация единиц O-антигена происходит в периплазматическом пространстве специализированной полимеразой Wzy, и полимерный фрагмент далее присоединяется к коровому липиду A лигазой WaaL, образуя липополисахарид[11][12]. Аппарат переноса молекул липополисахарида наружу клетки состоит из белков LptA, LptB, LptC, LptD, LptE. Для пяти из них удалось установить, в каких частях клетки они находятся, что может помочь разобраться в том, как функционирует аппарат сборки и выделения молекул липополисахарида[12]. Известно, что LptC переносит липополисахарид с клеточной мембраны во внешнюю[12]. LptE формирует комплекс с LptD, который обеспечивает встраивание молекул липополисахарида во внешнюю мембрану[12][13][14].
Везикулы наружной мембраны
От наружной мембраны могут отпочковываться везикулы (везикулы бактериальной внешней мембраны[en]*) диаметром от 20 до 500 нм. Образование везикул может быть связано с ростом бактериальной клетки, они могут служить средством доставки ферментов и прочих белков, например, патогенные бактерии могут транспортировать в составе везикул факторы вирулентности[en]. Например, у Pseudomonas aeruginosa в составе везикул наружной мембраны в числе прочих белков выделяется β-лактамаза, разрушающая пенициллин[15].
Примечания
- ↑ 1 2 Пиневич, 2006, с. 240.
- ↑ Пиневич, 2006, с. 241.
- ↑ 1 2 3 Пиневич, 2006, с. 250.
- ↑ Пиневич, 2006, с. 257.
- ↑ Пиневич, 2006, с. 240—241.
- ↑ Feldman M. F., Marolda C. L., Monteiro M. A., Perry M. B., Parodi A. J., Valvano M. A. The activity of a putative polyisoprenol-linked sugar translocase (Wzx) involved in Escherichia coli O antigen assembly is independent of the chemical structure of the O repeat. (англ.) // The Journal Of Biological Chemistry. — 1999. — 3 December (vol. 274, no. 49). — P. 35129—35138. — PMID 10574995. [исправить]
- ↑ Liu D., Cole R. A., Reeves P. R. An O-antigen processing function for Wzx (RfbX): a promising candidate for O-unit flippase. (англ.) // Journal Of Bacteriology. — 1996. — April (vol. 178, no. 7). — P. 2102—2107. — PMID 8606190. [исправить]
- ↑ Doerrler W. T., Reedy M. C., Raetz C. R. An Escherichia coli mutant defective in lipid export. (англ.) // The Journal Of Biological Chemistry. — 2001. — 13 April (vol. 276, no. 15). — P. 11461—11464. — DOI:10.1074/jbc.C100091200. — PMID 11278265. [исправить]
- ↑ Polissi A., Georgopoulos C. Mutational analysis and properties of the msbA gene of Escherichia coli, coding for an essential ABC family transporter. (англ.) // Molecular Microbiology. — 1996. — June (vol. 20, no. 6). — P. 1221—1233. — PMID 8809774. [исправить]
- ↑ Zhou Z., White K. A., Polissi A., Georgopoulos C., Raetz C. R. Function of Escherichia coli MsbA, an essential ABC family transporter, in lipid A and phospholipid biosynthesis. (англ.) // The Journal Of Biological Chemistry. — 1998. — 15 May (vol. 273, no. 20). — P. 12466—12475. — PMID 9575204. [исправить]
- ↑ Raetz C. R., Whitfield C. Lipopolysaccharide endotoxins. (англ.) // Annual Review Of Biochemistry. — 2002. — Vol. 71. — P. 635—700. — DOI:10.1146/annurev.biochem.71.110601.135414. — PMID 12045108. [исправить]
- ↑ 1 2 3 4 Sperandeo P., Lau F. K., Carpentieri A., De Castro C., Molinaro A., Dehò G., Silhavy T. J., Polissi A. Functional analysis of the protein machinery required for transport of lipopolysaccharide to the outer membrane of Escherichia coli. (англ.) // Journal Of Bacteriology. — 2008. — July (vol. 190, no. 13). — P. 4460—4469. — DOI:10.1128/JB.00270-08. — PMID 18424520. [исправить]
- ↑ Wu T., McCandlish A. C., Gronenberg L. S., Chng S. S., Silhavy T. J., Kahne D. Identification of a protein complex that assembles lipopolysaccharide in the outer membrane of Escherichia coli. (англ.) // Proceedings Of The National Academy Of Sciences Of The United States Of America. — 2006. — 1 August (vol. 103, no. 31). — P. 11754—11759. — DOI:10.1073/pnas.0604744103. — PMID 16861298. [исправить]
- ↑ Bos M. P., Tefsen B., Geurtsen J., Tommassen J. Identification of an outer membrane protein required for the transport of lipopolysaccharide to the bacterial cell surface. (англ.) // Proceedings Of The National Academy Of Sciences Of The United States Of America. — 2004. — 22 June (vol. 101, no. 25). — P. 9417—9422. — DOI:10.1073/pnas.0402340101. — PMID 15192148. [исправить]
- ↑ Пиневич, 2006, с. 256.
Литература
- Пиневич А. В. Микробиология. Биология прокариотов: в 3 т. — СПб.: Издательство С.-Петербургского университета, 2006. — Т. I. — 352 с. — ISBN 5-288-04057-5.
wiki2.org
Внешняя бактериальная мембрана — WiKi
Состав
Строение липополисахаридаВнешняя мембрана, как и клеточная, имеет толщину 7—8 нм[1]. Подобно клеточной мембране, внешняя мембрана представляет собой липидный бислой, который состоит из фосфолипидов, преимущественно фосфатидилэтаноламина, фосфатидилглицерола[en] и дифосфатидилглицерола. Однако во внешней мембране фосфолипиды распределены между слоями несимметрично: внешний лист мембраны образован преимущественно липополисахаридами. Исключениями являются цианобактерии и виды рода Neisseria, у которых во внешнем слое мембраны одновременно присутствуют фосфолипиды и липоолигосахариды. В структуре липополисахарида можно выделить четыре структурных блока:
Внешняя мембрана богата белками, их обозначают Omp[en] (от англ. outer membrane protein). Белки могут покрывать до половины поверхности внешней мембраны. Белки Omp подразделяют на главные и минорные. Главные Omp-белки синтезируются постоянно; к их числу относятся липопротеин Брауна, липопротеины спирохет, OmpA и неспецифический порин[en] OmpF[3].
В структурно-функциональном отношении все белки внешней мембраны можно подразделить на три группы:
Иногда на препаратах обнаруживаются зоны контакта клеточной и внешней мембран. Такие участки называют контактами Байера. Вероятно, непосредственного контакта между мембранами не происходят, и они связываются друг с другом специальными белками в образующейся на месте контакте бреши в пептидогликановом слое[4].
Функции
Во внешней мембране имеются постоянно открытые ионные каналы, из-за чего на ней не может поддерживаться постоянный электрохимический градиент ионов натрия или протонов, поэтому внешняя мембрана не принимает участия в метаболизме клетки. Она также не имеет отношения к биосинтезу белков, липидов и полисахаридов, хотя может быть задействована в их секреции. Однако во внешней мембране находятся некоторые ферменты — пермеазы, гидролазы, иногда — окислительные ферменты[en], такие как марганец-оксидаза[1].
Функции внешней мембраны в бактериальной клетке очень разнообразны. Она совместно с клеточной мембраной образует периплазматическое пространство (периплазму), наряду с клеточной стенкой придаёт жёсткость клетке, служит фильтром, который не даёт попасть в периплазму крупным гидрофильным молекулам и задерживает гидрофобные молекулы. Она не даёт покинуть клетку многим ферментам периплазмы, участвует в поступлении в клетку питательных веществ, а также выделении наружу антибиотиков, токсинов, метаболитов и разнообразных белков. Внешняя мембрана опосредует неспецифичную адгезию бактериальных клеток, взаимодействует с бактериофагами, поверхностными рецепторами как прокариотических, так и эукариотических клеток, а также антителами. Благодаря внешней мембране жёлчные кислоты и другие амфифильные детергенты, а также антибиотики действуют на грамотрицательные бактерии слабее, чем на грамположительные[5]. Липоолигосахариды — важнейшие антигены патогенных грамотрицательных бактерий, их также называют эндотоксинами[3].
Биогенез
Механизмы, которые обеспечивают доставку компонентов внешней мембраны на поверхность клетки, до конца не ясны. Компоненты липополисахарида — липид A и O-антигеновые повторяющиеся единицы — синтезируются на цитоплазматической стороне клеточной мембраны и доставляются наружу независимо двумя специализированными транспортными системами, а именно, переносчиком O-антигена Wzx (RfbX) и ABC-транспортера[en] MsbA, который перемещает липид A с внутреннего липидного слоя клеточной мембраны в наружный[6][7][8][9][10]. Полимеризация единиц O-антигена происходит в периплазматическом пространстве специализированной полимеразой Wzy, и полимерный фрагмент далее присоединяется к коровому липиду A лигазой WaaL, образуя липополисахарид[11][12]. Аппарат переноса молекул липополисахарида наружу клетки состоит из белков LptA, LptB, LptC, LptD, LptE. Для пяти из них удалось установить, в каких частях клетки они находятся, что может помочь разобраться в том, как функционирует аппарат сборки и выделения молекул липополисахарида[12]. Известно, что LptC переносит липополисахарид с клеточной мембраны во внешнюю[12]. LptE формирует комплекс с LptD, который обеспечивает встраивание молекул липополисахарида во внешнюю мембрану[12][13][14].
Везикулы наружной мембраны
Примечания
- ↑ 1 2 Пиневич, 2006, с. 240.
- ↑ Пиневич, 2006, с. 241.
- ↑ 1 2 3 Пиневич, 2006, с. 250.
- ↑ Пиневич, 2006, с. 257.
- ↑ Пиневич, 2006, с. 240—241.
- ↑ Feldman M. F., Marolda C. L., Monteiro M. A., Perry M. B., Parodi A. J., Valvano M. A. The activity of a putative polyisoprenol-linked sugar translocase (Wzx) involved in Escherichia coli O antigen assembly is independent of the chemical structure of the O repeat. (англ.) // The Journal Of Biological Chemistry. — 1999. — 3 December (vol. 274, no. 49). — P. 35129—35138. — PMID 10574995. [исправить]
- ↑ Liu D., Cole R. A., Reeves P. R. An O-antigen processing function for Wzx (RfbX): a promising candidate for O-unit flippase. (англ.) // Journal Of Bacteriology. — 1996. — April (vol. 178, no. 7). — P. 2102—2107. — PMID 8606190. [исправить]
- ↑ Doerrler W. T., Reedy M. C., Raetz C. R. An Escherichia coli mutant defective in lipid export. (англ.) // The Journal Of Biological Chemistry. — 2001. — 13 April (vol. 276, no. 15). — P. 11461—11464. — DOI:10.1074/jbc.C100091200. — PMID 11278265. [исправить]
- ↑ Polissi A., Georgopoulos C. Mutational analysis and properties of the msbA gene of Escherichia coli, coding for an essential ABC family transporter. (англ.) // Molecular Microbiology. — 1996. — June (vol. 20, no. 6). — P. 1221—1233. — PMID 8809774. [исправить]
- ↑ Zhou Z., White K. A., Polissi A., Georgopoulos C., Raetz C. R. Function of Escherichia coli MsbA, an essential ABC family transporter, in lipid A and phospholipid biosynthesis. (англ.) // The Journal Of Biological Chemistry. — 1998. — 15 May (vol. 273, no. 20). — P. 12466—12475. — PMID 9575204. [исправить]
- ↑ Raetz C. R., Whitfield C. Lipopolysaccharide endotoxins. (англ.) // Annual Review Of Biochemistry. — 2002. — Vol. 71. — P. 635—700. — DOI:10.1146/annurev.biochem.71.110601.135414. — PMID 12045108. [исправить]
- ↑ 1 2 3 4 Sperandeo P., Lau F. K., Carpentieri A., De Castro C., Molinaro A., Dehò G., Silhavy T. J., Polissi A. Functional analysis of the protein machinery required for transport of lipopolysaccharide to the outer membrane of Escherichia coli. (англ.) // Journal Of Bacteriology. — 2008. — July (vol. 190, no. 13). — P. 4460—4469. — DOI:10.1128/JB.00270-08. — PMID 18424520. [исправить]
- ↑ Wu T., McCandlish A. C., Gronenberg L. S., Chng S. S., Silhavy T. J., Kahne D. Identification of a protein complex that assembles lipopolysaccharide in the outer membrane of Escherichia coli. (англ.) // Proceedings Of The National Academy Of Sciences Of The United States Of America. — 2006. — 1 August (vol. 103, no. 31). — P. 11754—11759. — DOI:10.1073/pnas.0604744103. — PMID 16861298. [исправить]
- ↑ Bos M. P., Tefsen B., Geurtsen J., Tommassen J. Identification of an outer membrane protein required for the transport of lipopolysaccharide to the bacterial cell surface. (англ.) // Proceedings Of The National Academy Of Sciences Of The United States Of America. — 2004. — 22 June (vol. 101, no. 25). — P. 9417—9422. — DOI:10.1073/pnas.0402340101. — PMID 15192148. [исправить]
- ↑ Пиневич, 2006, с. 256.
Литература
- Пиневич А. В. Микробиология. Биология прокариотов: в 3 т. — СПб.: Издательство С.-Петербургского университета, 2006. — Т. I. — 352 с. — ISBN 5-288-04057-5.
ru-wiki.org
функции, строение. Наружная цитоплазматическая мембрана :: SYL.ru
Наружная цитоплазматическая мембрана представляет собой тончайшую пленку. Ее толщина – порядка 7-10 нм. Просматривается пленка только в электронный микроскоп.  Далее рассмотрим, что собой представляет цитоплазматическая мембрана. Функции пленки также будут описаны в статье.
Далее рассмотрим, что собой представляет цитоплазматическая мембрана. Функции пленки также будут описаны в статье.
Структура
Какой состав имеет цитоплазматическая мембрана? Строение пленки достаточно разнообразно. В соответствии с химической организацией, она представляет собой комплекс белков и липидов. Цитоплазматическая мембрана клетки включает в себя бислой. Он выступает в качестве основы. Кроме этого, цитоплазматическая мембрана содержит холестерол и гликолипиды. Этим веществам свойственна амфипатричность. Другими словами, в них присутствуют гидрофобные (“боящиеся влаги”) и гидрофильные (“любящие воду”) концы. Последние (фосфатная группа) направлены наружу от мембраны, вторые (остатки от жирных кислот) ориентированы друг к другу. За счет этого и формируется липидный биполярный слой. Липидные молекулы обладают подвижностью. Они способны перемещаться в собственном монослое либо (что редко) из одного в другой.  Липидный слой может иметь состояние твердого или жидкого кристалла. Монослои отличаются асимметричностью. Это значит, что в них различен состав липидов. За счет этого свойства цитоплазматические мембраны обладают специфичностью даже в рамках одной клетки. Ко второму обязательному компоненту пленки относят белки. Многие из этих соединений могут перемещаться в мембранной плоскости либо совершать вращения вокруг собственной оси. При этом они не способны переходить из одной части бислоя в другую. Защита внутренней среды – основная задача, которую выполняет цитоплазматическая мембрана. Строение пленки, кроме этого, обеспечивает течение различных процессов. За выполнение тех или иных задач отвечают белки. Благодаря липидам обеспечиваются структурные особенности пленки.
Липидный слой может иметь состояние твердого или жидкого кристалла. Монослои отличаются асимметричностью. Это значит, что в них различен состав липидов. За счет этого свойства цитоплазматические мембраны обладают специфичностью даже в рамках одной клетки. Ко второму обязательному компоненту пленки относят белки. Многие из этих соединений могут перемещаться в мембранной плоскости либо совершать вращения вокруг собственной оси. При этом они не способны переходить из одной части бислоя в другую. Защита внутренней среды – основная задача, которую выполняет цитоплазматическая мембрана. Строение пленки, кроме этого, обеспечивает течение различных процессов. За выполнение тех или иных задач отвечают белки. Благодаря липидам обеспечиваются структурные особенности пленки.
Цитоплазматическая мембрана: функции
Основными задачами являются:
- Барьерная. Защитная пленка обеспечивает активный, пассивный, избирательный, регулируемый обмен соединений с внешней средой. За счет избирательной проницаемости осуществляется отделение клетки и ее компартментов и снабжение их нужными веществами.
- Транспортная. Сквозь пленку осуществляется переход соединений от клетки к клетке. Благодаря этому доставляются питательные соединения, удаляются конечные продукты обмена, происходит секреция разных веществ. Кроме этого, формируются ионные градиенты, на оптимальном уровне поддерживаются ионная концентрация и рН. Они необходимы для активной деятельности ферментов клетки.
Вспомогательные задачи
- Матричная. Эта функция обеспечивает определенную ориентацию и взаиморасположение белков мембраны, а также оптимальное их взаимодействие.
- Механическая. За счет нее обеспечивается автономность клетки, внутренних структур. Также осуществляется соединение элемента с прочими аналогичными.

- Энергетическая. На фоне фотосинтеза в хлоропластах и при осуществлении клеточного дыхания в мембранах активны системы энергетического переноса. В них также участвуют и белковые соединения.
- Рецепторная. Ряд белков, которые присутствуют в мембране, обеспечивает восприятие различных сигналов. К примеру, циркулирующие в крови стероиды оказывают воздействие только на те клетки-мишени, которые обладают соответствующими гормонам рецепторами. Химические соединения, обеспечивающие проведение импульсов (нейромедиаторы), также связываются с помощью особых белков клеток-мишеней.
Особые свойства
К специфическим функциям мембраны относят:
- Ферментативную. Зачастую белки, которые содержит цитоплазматическая мембрана, выступают в качестве ферментов.
- Генерацию и проведение биопотенциалов.
- Маркировку. Цитоплазматическая мембрана включает в свой состав особые антигены. Они действуют как маркеры-“ярлыки”. Благодаря им осуществляется распознание клеток. Маркеры представляют собой гликопротеины – белки, содержащие разветвленные олигосахаридные боковые цепи. Они выступают в качестве “антенн”. Благодаря огромному количеству вариантов боковых цепей для того или другого типа клеток может быть сформирован особый маркер. При их помощи распознанные друг другом элементы начинают действовать согласованно. К примеру, так происходит при образовании тканей и органов. Маркировка также позволяет иммунитету определить чужеродные антигены.
Дополнительные сведения
Если какие-то частицы по тем или другим причинам не способны пройти сквозь фосфолипидный бислой (к примеру, вследствие гидрофильных свойств, поскольку внутри цитоплазматическая мембрана гидрофобна и такие соединения не пропускает, либо из-за больших размеров самих частиц), но они необходимы, то пройти они могут с помощью специальных белков-переносчиков (транспортеров) и белков-каналов. Либо проникновение их осуществляется посредством эндоцитоза. В процессе пассивного транспорта пересечение веществами липидного слоя происходит путем диффузии. При этом энергия не затрачивается. В качестве одного из вариантов такого механизма может выступать облегченная диффузия. В ходе нее облегчает прохождение вещества какая-нибудь специфическая молекула. У нее может присутствовать канал, способный пропускать только однотипные частицы. При активном транспорте затрачивается энергия. Это связано с тем, что данный процесс осуществляется против концентрационного градиента. Цитоплазматическая мембрана содержит особые белки-насосы, АТФазу в том числе, которая способствует активному вхождению калиевых и выведению натриевых ионов.
Модели
Их существует несколько:
- “Бутербродная модель”. Идею о трехслойном строении всех мембран высказали ученые Даусон и Даниэли в 1935 году. По их мнению, структура пленки была следующей: белки-липиды-белки. Такое представление существовало достаточно долго.
- “Жидкостно-мозаичная структура”. Эта модель была описана Николсоном и Сингером в 1972 году. В соответствии с ней белковые молекулы не формируют сплошной слой, а погружаются в биполярный липидный в виде мозаики на различную глубину. Эта модель считается наиболее универсальной.
- “Белково-кристаллическая структура”. В соответствии с этой моделью мембраны формируются за счет переплетения белковых и липидных молекул, которые объединены на базе гидрофильно-гидрофобных связей.
www.syl.ru